
1) Adkrustacja, apozycja (powlekanie) - jest to odkładanie związków na powierzchni (wewnętrznej lub zewnętrznej) ściany komórkowej pierwotnej u roślin. Substancje powlekające to głównie kutyna, woski, suberyna (związki o tłuszczowym charakterze), a także kaloza i śluzy. Adkrustacja jest ochroną przed nadmiernym przepuszczaniem wody przez ścianę. Niekiedy rozrastająca się wtórna ściana komórkowa jest tak gruba, że zajmuje prawie całą objętość komórki, na przykład we włóknach sklerenchymatycznych. Natomiast w komórkach merystematycznych i miękiszowych ściana komórkowa jest cienka i cały czas zachowuje pierwotny charakter. Jeżeli adkrustacja kutyną dotyczy zewnętrznych ścian komórek stanowiących zewnętrzną warstwę rośliny wówczas proces taki nazywany jest kutykularyzacją. Wewnętrzna powierzchnia ściany komórkowej adkrustowana jest suberyną w trakcie procesu suberynizacji (korkowacenia). Powierzchni ścian ziaren pyłku oraz zarodników mszaków, paprotników, glonów oraz grzybów pokryta jest mieszaniną związków chemicznych określaną jako sporopolenina.
2) Aksenia - jest to bierna odporność roślin na patogeny, jest odpornością konstytutywną, czyli istniejącą niezależnie od kontaktu z patogenem. Formą aksenii są przystosowania anatomiczno-morfologiczne zapewniające barierę przed wniknięciem czynników chorobotwórczych. Barierami takimi są kutykula lub woskowy nalot pokrywający liście oraz nieliczne i małe aparaty szparkowe. Aksenia obejmuje także przystosowania fizjologiczno-chemiczne istniejące w roślinie przez cały czas.
3) Allomony (z gr. allos - inny) - są to substancje semiochemiczne (substancje semiochemiczne to związki chemiczne używane przez zwierzęta oraz rośliny do przekazywania informacji, wywierania określonego wpływu na otoczenie, głównie w celach obronnych, wszczynania alarmu, zaznaczania własnego terytorium) wywołujące reakcję korzystną jedynie dla dawcy. Należą do nich antybiotyki, repelenty czy substancje zwabiające ofiary. Przykładem allomonu jest proherbicyd - 1, 4, 5 -trihydroksynaftalen, wytwarzany przez liście orzecha włoskiego, który po spłukaniu deszczem opada na ziemię, gdzie zostaje utleniony do 5-hydroksynaftochinonu- 1,4, związku hamującego wzrost niektórych roślin (działanie allelopatyczne). Do allomonów zalicza się również kocineliny, alkaloidy wytwarzane przez biedronki, które odstraszają drapieżniki chętne do konsumowania tychże chrząszczy. Termin allomony wprowadzony został w 1968 roku przez W.L Browna i T. Eisnera dla określenia międzygatunkowych sygnałów chemicznych. Później wprowadzono podział tych sygnałów na allomony w wyżej przedstawionym znaczeniu oraz kairomony.
4) Apoplast - jest to zespół ścian komórkowych, wnętrz martwych komórek i przestworów międzykomórkowych w organizmie roślinnym. Połączone ze sobą i wyscycone wodę ściany komórkowe razem z wypełnionymi wodą wnętrzami martwych komórek przewodzącycch są głównymi drogami transportu wody w roślinie. W ścianach komórkowych ruch wody możliwy jest dzięki zjawisku imbibicji. Wypełnione powietrzem przestwory międzykomórkowe stanowią rezerwuary gazów i umożliwiają roślinie prowadzenie wymiany gazowej z otoczeniem.
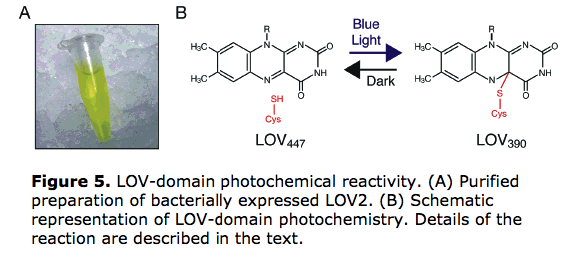
5) Białka LEA (ang. Late-embryogenesis abundant proteins) - jest to grupa silnie hydrofilowych białek wytwarzanych podczas późnej embriogenezy roślin i zapewniających odporność na wysuszenie nasionom. Białka są wytwarzane przez rośliny także podczas stresu wodnego. Po raz pierwszy zostały wyizolowane z nasion bawełny i opisane w roku 1981 przez Leona Dura. Białka LEA należą do większej grupy hydrofilin. Na podstawie podobieństw sekwencji dzielone są na kilka rodzin. Wspólną cechą poza wysoką hydrofilowością jest wysoka zawartość glicyny oraz alaniny i seryny. Nie stwierdza się w nich obecności reszty cysteinowych i tryptofanowych. Zostały wykryte u wielu grup roślin oraz w grzybach, nicieniach, bakteriach i archeowcach. Do ich zwiększonej syntezy dochodzi w roślinach podczas suszy oraz mrozów. Chociaż udział białek LEA w odpowiedzi na stres wodny i niską temperaturę jest powszechnie uznawany, to sam mechanizm ochronny jest nieznany. Na podstawie wyników badań rozważana jest ich funkcja jako przeciwutleniacza, substancji stabilizujących błony i białka oraz pośrednio udział w ochronie komórki jako tarcze molekularne. Rozważana jest także funkcja "wypełniacza przestrzeni" chroniącego komórkę przed uszkodzeniem struktur przy silnym ograniczeniu objętości. Szczególną rolę wydają się spełniać w ochronie błon mitochondrialnych przy silnym odwodnieniu komórki.
6) Białka szoku cieplnego (HSP - ang. Heat shock proteins) - jest to grupa białek, których ekspresja wzrasta, kiedy komórki są narażone na działanie czynników stresowych, m.in. podwyższonej temperatury, ale również niskiej temperatury, stresu solnego, osmotycznego i metali ciężkich. Produkcja HSP może wzrastać także w odpowiedzi na infekcje, zapalenie, działanie toksyn, promieniowanie UV, głodzenie, niedotlenienie itp. Część HSP jest produkowana w komórce cały czas. HSP działają jako białka opiekuńcze odpowiedzialne za prawidłowe zwijanie się innych białek, ich ilogomeryzacją, translokację oraz degradację. Funkcje HSP są podobne u wszystkich organizmów żywych. Ich sekwencja aminokwasowa jest silnie konserwatywna. Występują zarówno u organizmów prokariotycznych jak i eukariotycznych. Po raz piewszy zostały odkryte u muszki owocowej. Ze względu na masę cząsteczkową (wyrażona w kDa) białka szoku cieplnego dzieli się na pięć głównych rodzin: niskocząsteczkowe hsp60, hsp70, hsp90 i hsp100.








30) Kryptochromy (z gr. krupto chroma - ukryty kolor) - są to fotoreceptory światła niebieskiego u zwierząt i roślin. Nazwy tej używa się w odniesieniu do specyficznej podklasy receptorów światła niebieskiego - flawoprotein regulujących kiełkowanie, elongację, fotoperiodyzm i inne reakcje wzrostowe roślin. Światło niebieskie pośredniczy także w fototropizmie, ale udział kryptochromów w tym procesie nie jest znany. W tym procesie bierze udział NPH1, białko zawierające flawinową grupę chromoforową, którego widmo absorpcyjne funkcjonalnej cząsteczki pokrywa się z widmem czynnościowym dla procesu fototropizmu i nazywane jest fototropiną. Kryptochromy to białka wczesne ewolucyjnie, pochodzące od fotoliaz - bakteryjnych enzymów flawoproteinowych aktywowanych przez światło i uczestniczących w naprawie dimerów pirymidynowych DNA. U eukariontów kryptochromy utraciły tę zdolność katalityczną. Kryptochromy odkryto również u koralowców, gdzie biorą udział w koordynacji cyklów rozmnażania w czasie kilku wiosennych nocy podczas pełni księżyca. Znaleziono je również u owadów i ssaków. Dwa kryptochromy znalezione u ssaków odgrywają zasadniczą rolę w rozmnażaniu oraz utrzymywaniu rytmu dobowego. Badania sugerują, że kryptochromy ułatwiają nawigację ptaków podczas ich migracji, ponieważ są podstawą funkcjonowania ich magnetycznego zmysłu używanego do wyczuwania pola magnetycznego Ziemi. Inne teorie sugerują, że te zdolności są obecne także u ssaków, ale są uśpione. W ludzkim genomie, geny kodujące Cry1 i Cry2 zostały znalezione odpowiednio na chromosomach 12 i 11.
37) Morfaktyny - jest to grupa syntetycznych regulatorów wzrostu i rozwoju roślin, których działanie związane jest z zahamowaniem transportu auksyn. Wpływ pochodnych fluorenu na morfogenezę roślin zaobserwowano w latach 60. XX wieku. Działanie morfaktyn polega na ogólnym zahamowaniu wzrostu roślin. Zahamowaniu ulega kiełkowanie nasion. U roślin zarówno jednoliściennych jak i dwuliściennych potraktowanych pochodnymi fluorenu obserwowano zanik reakcji fototropicznych oraz grawitropicznych w wierzchołkach wzrostu pędu i korzenia. Przykłady: Chloroflurenol, Flurenol, Methyl benzilate, Methyl chloroflurenol, Methyl dichloroflurenol. Sam fluoren jest nieaktywny, ale sama addycja grupy COOH w pozycji 9' powoduje jego aktywację.

4) Apoplast - jest to zespół ścian komórkowych, wnętrz martwych komórek i przestworów międzykomórkowych w organizmie roślinnym. Połączone ze sobą i wyscycone wodę ściany komórkowe razem z wypełnionymi wodą wnętrzami martwych komórek przewodzącycch są głównymi drogami transportu wody w roślinie. W ścianach komórkowych ruch wody możliwy jest dzięki zjawisku imbibicji. Wypełnione powietrzem przestwory międzykomórkowe stanowią rezerwuary gazów i umożliwiają roślinie prowadzenie wymiany gazowej z otoczeniem.
 |
| Źródło: mpimp-golm.mpg.de - Schemat działania białek LEA. Integracja z błoną komórkową. |
 |
| Źródło: Wikipedia. Model cząsteczki białka GroES/ GroEL. |
 |
| Źródło: Wikipedia. Hsp90. |
- HSP90 (z ang. heat shock protein 90) - to białko z rodziny białek szoku cieplnego, należące do białek chaperonowych (opiekuńczych). Hsp90 występuje u eubakterii i wszystkich eukariontów, ale wydaje się być nieobecne w komórkach archeowców. Cytoplazmatyczne Hsp90 jest niezbędne do prawidłowego funkcjonowania komórek eukariontów w każdej sytuacji, natomiast u bakterii produkt białkowy bakteryjnego homologu genu HtpG jest zbędny w warunkach innych aniżelis zok cieplny. Na molekularnym poziomie białko Hsp90 odpowiada za pomoc w zwijaniu innych białek, m.in. stąd wynika jego ochronna rola polegająca na stabilizacji białek w czasie szoku cieplnego. Hsp90 oddziałuje z ponad 10% białek, wliczając w to białka związane z procesem nowotworzenia, np. białkiem p53. W związku z tym, inhibitory białka Hsp90, takie jak geldanamycyna i jej analogiii, są intensywnie badanie jako leki przeciwnowotworowe. Białka szoku cieplneog, wliczając w to Hsp90, są jednymi z najintensywniej produkowanych białek (ok. 1-2% wszystkich białek; dodatkowo w warunkach stresu, gdy ulegają intensywnej produkcji, ich ilość może wynosić 4-6%). Ponadto, białka te są wysoko konserwowane ewolucyjnie (60% identyczności sekwencyjnej między drożdżowym i ludzkim Hsp90alfa).
- >>IZOFORMY: U ssaków, obecne są przynajmniej dwa geny kodujące cytoplazmatyczne formy białka Hsp90 (identyczność na poziomie sekwencji dla Hsp90alfa i Hsp90beta wynosi 85%). Obie formy powstały najprawdopodobniej w wyniku duplikacji genów wiele milionów lat temu. W sumie u ludzi jest 5 funkcjonalnych genów kodujących następujące izoformy:

Ponadto, istnieje 12 pseudogenów Hsp90 - genów, które nie ulegają ekspresji. Dodatkowo, wykryto trzeci wariant cytoplazmatyczny Hsp90N (pozbawiony domeny wiążącej ATP). Jednak, po bardziej szczegółowej analizie okazało się, że izoforma ta nie istnieje i jest artefaktem klonowania.
7) Chemotyp - jest to fenotyp chemiczny, wyrównany nie ze względu na obecność lub stężenie specyficznych metabolitów wtórnych. Czynniki genetyczne i epgienetyczne mogą nie wiązać się z określonymi cechami morfologicznymi - morfotypem lub anatomicznymi lecz powodować wystąpienie unikalnych cech chemicznych organizmu. Pojęcie zostało zastosowane po raz pierwszy przez Rolfa Santessona i jego syna Johana w roku 1968 w odniesieniu do roślin. Współcześnie termin stosowany jest także w odniesieniu do mikroorganizmów.
Wyróżnianie chemotypów roślin ma znaczenie w farmacji, przy wykorzystywaniu substancji aktywnych pochodzenia roślinnego. Obecność i stężenie takiej substancji decyduje o właściwościach leczniczych preparatów ziołowych. Określenie chemotypu jest również wykorzystywane w badaniach taksonomicznych. Skład chemiczny może być podstawą wyróżnienia odrębnych taksonów. Analiza zawartości kannabinoidów w Cannabis potwierdza kocencpję wyróżniania gatunku C. sativa i C. indica. Ze względu na wykorzystanie kannabinoidy w przemyśle farmaceutycznym chemotypy o wysokiej zawartości Δ (9)-tetrahydrokanabinolu i kanabidiolu są przedmiotem zainteresowania koncernów farmaceutycznych. Poszukiwanie odpowiednich chemotypów może mieć na celu pozyskanie substancji zapachowych wykorzystywanych przez człowieka. Wyróżnianie chemotypów jest ważne także ze względu na obronę roślin przed szkodnikami. Różne profile chemiczne mogą skutkować zróżnicowaniem wrażliwości upraw względem szkodników. Substancja pozwalająca wyróżnić chemotyp może być syntetyzowana u roślin po odebraniu sygnału o obecności roślinożercy.
8) Chlorooddychanie (chlororespiracja) - jest to zjawisko podobne do łańcucha oddechowego, jednakże zachodzące w chloroplastach. W chloroplastach glonów i roślin wyższych możliwa jest niefotochemiczna redukcja puli plastochinonu ze zużyciem NADH lub NADPH. Plastochinon może być następnie utleniany przez mało znaną oksydazę. W efekcie zachodzenia chloroodychania chloroplasty mogą zużywać NADPH i wytwarzać gradient protonowy wykorzystywany do syntezy ATP. Natężenie procesu redukcji plastochinonu bez udziału światła jest niewielkie, jego rola nie jest znana, a samo zjawisko jest zbadane w niewielkim stopniu. Po raz pierwszy dowody na zachodzenie niefotochemicznej redukcji plastochinonu przedstawiono w 1982 roku. Znaczenie chlorooddychania wyjaśniono jako mechanizm zapewniający przetwarzanie ATP i NADPH generowanego w procesie glikolizy. W dalszych badaniach stwierdzono występowanie w błonach tylakoidów kompleksu dehydrogenazy NAD(P)H - Ndh zbliżonego do kompleksu dehydrogenazy NADH obecnego w mitochondriach. Kompleks Ndh zawarty w chloroplastach kodowany jest przez genom chloroplastowy. Istnienie drugiego składnika biorącego udział w chloroddychaniu - oksydazy - udało się potwierdzić w chloroplastach Arabidopsis thaliana. Enzym ten jest kodowany przez genom jądrowy i ma strukturę podobną do mitochondrialnej oksydazy alternatywnej.
9) Cybrydy
10) Cykl glioksylanowy
11) Dekarboksylaza glicyny (Kompleks enzymatyczny dekarboksylazy glicyny - GDC, EC, 2.1.2.10) - jest to kompleks enzymatyczny, który razem z hydroksymetylotransferazą seryny odpowiedzialny jest za przekształcenie glicyny do seryny w mitochondriach roślinnych w procesie fotooddychania. Dekarboksylaza glicyny składa się z czterech różnych komponentów enzymatycznych: dwóch podjednostek P (dimer o masie 98-106 kDa), 27 podjednostek H (monomer o masie 13,8-15,5 kDa), 9 podjednostek T (monomer o masie 41-45 kDa) i dwóch podjednostek L (monomer o masie 50-59 kDa). Białko H nie pełni funkcji katalitycznych. Jest zlokalizowane w centrum kompleksu i przenosi produkty pośrednie między pozostałymi trzema komponentami. Białko P rozpoczyna reakcję dekarboksylacji poprzez przyłączenie grupy alfa-aminowej glicyny do 5-fosforanu pirydoksalu. Następuje przeniesienie elektronów a atomu węgla alfa i odłączenie grupy karboksylowej. Pozostałe elementy cząsteczki glicyny: węgiel alfa i grupa alfa-aminowa przenoszone są poprzez podjednostkę H do centrum aktywnego białka T. Białko T katalizuje reakcję przyłączenia atomu węgla z grupy metyloaminowej na tetrahydrofolian. Jednocześnie odłączana jest cząsteczka NH3. Oderwane elektrony oraz para atomów wodoru w postaci kwasu dihydroliponowego przenoszone są na podjednostkę L i służą do redukcji FAD do FADH2. Siłą redukcyjna z FADH2 jest następnie przenoszona na cząsteczkę NAD+. Ostatecznie uwalnana jest cząsteczka CO2, NH3 oraz powstaje cząsteczka NADH. Grupa metylowa przyłączana jest przy udziale SHMT do cząsteczki glicyny co prowadzi do powstania seryny. SHMT jest tetramerem o masie 220 kDa składającym się z czterech identycznych podjednostek o masie 53 kDa. Pewną aktywność dekarboksylazy glicyny wykazują prawdopodobnie wszystkie komórki roślinne. Największą aktywność enzym ten wykazuje w fotosyntetyzujących tkankach roślin typu C3. W mitochondriach grochu GDC stanowi 1/3 rozpuszczalnych białek mitochondrialnych. W tkankach niefotosyntetyzujących i roślinach etiolowanych aktywność GDC jest znikoma.
12) Dojrzałość konsumpcyjna:
 |
| Źródło: Wikipedia. |
13) Dojrzałość zbiorcza owoców:
 |
| Źródło: Wikipedia. |
14) Ekstensyny - rodzina glikoprotein bogatych w hydroksyprolinę, budujących strukturę ściany komórek roślin. Masa monomerów ekstensyny została początkowo oszacowana na 80 kDa. W kolejnych badaniach natywny prekursor ekstensyn miała masę 240-300 kDa. Do licznych reszt hydroksyproliny przyłączona jest L-arabinoza. Ściana komórkowa roślin jest kompozytem składającym się z trzech wzajemnie przenikających się sieci. Dwie z tych sieci zbudowane są z polisacharydów a trzecie z glikoprotein. W genomie Arabidopsis thaliana kodowanych jest 20 ściśle homologicznych polipeptydów ekstensyn. Polipeptydy te są niezbędne do wykształcenia ściany komórkowej podczas cytokinezy. Potwierdzają to obserwowane zaburzenia na etapie embriogenezy u mutanta niezdolnego do syntezy ekstensyny. Podwyższona zawartość ekstensyn umożliwia ochronę tkanek roślinnych przed patogenami. Usieciowanie powstające w wyniku działania peroksydaz i H2O2 pozwala uzyskać barierę przez którą nie są w stanie przeniknąć mikroorganizmy.
 |
| Źródło: itqb.uni.pt - Hipoteza działania ekstensyn. |
15) Eksudat - wydzielina z korzeni roślin, służąca allelopatii lub pobieraniu jonów z gleby.
16) Etiolacja (wypłonienie) - zjawisko polegające na niewytworzeniu zielonego barwnika chlorofilu w tkankach oraz szybkim wzroście wydłużeniowym łodygi roślin rozwijających się w ciemności. Podczas wzrostu w ciemności roślina wykorzystuje materiały zapasowe. Gdy znajdzie się na świetle, zachodzi proces deetiolacji, polegający na zahamowaniu wzrostu wydłużeniowego łodygi, rozwinięciu liści, wytworzeniu chlorofilu i rozpoczęciu procesu fotosyntezy. Receptorem światła w procesie deetiolacji jest fitochrom, który w etiolowanych siewkach występuje w dużych ilościach. Naświetlenie takich siewek światłem czerwonym lub białym. Powstający w wyniku oświetlenia fitochrom PFR występuje jako Phy A i Phy B. Forma A jest nietrwała i ulega szybkiemu rozpadowi. Rozpad fitochromu na świetle jest zjawiskiem niedowracalnym, a wyjatkowo wysokie stężenie występuje tylko u siewek.
 |
| Źródło: Wikipedia. Białe szparagi. |
 |
| Źródło: Wikipedia. Struktura Fitochelatyny. |
17) Fitochelatyny - są to polipeptydy wytwarzane w komórkach roślinnych i mikroorganizmach, uczestniczące w wychwytywaniu jonów toksycznych metali. Po połączeniu z metalem kompleks fitochelatyna-metal transportowany jest do wakuoli. W ten sposób cytoplazma poddawana jest detoksykacji. PC zostały wykryte i opisane w roku 1981 u drożdży. W roku 1985 wykryto polipeptyd wiążący metale ciężkie u roślin wyższych, wtedy również zaczęto stosować nazwę fitochelatyny. W roku 1985 został zidentyfikowany enzym odpowiedzialny z ich syntezę, syntaza fitochelatyn. Badania genetyczne wykazały, że polipeptydy podobne do tych odkrytych u roślin mogą być wytwarzane w organizmach niektórych zwierząt. Prekursorem fitochelatyn jest powszechnie występującym w organizmach roślinnym tripeptyd glutation. Charakterystycznym elementem fitochelatyn jest wielokrotnie powtórzone połączenie reszty kwasu glutaminowego (Glu) i cysteiny (Cys). W skład polipeptydu wchodzi także trzeci aminokwas, którym może być glicyna (Gly), alanina (Ala) lub glutamina (Gln). Możliwe są różne warianty peptydu (Glu-Cys)n-Gly (fitochelatyna) oraz (Glu-Cys)n-Ala (homofitochelatyna), (Glu-Cys)n-Ser (hydroksymetylofitochelatyna), (Glu-Cys)n-Glu (izo-fitochelatyna (Gln))), (Glu-Cys)n (des-Gly-fitochelatyna), gdzie n wynosi od 2 do 11. Fitochelatyny są klasyfikowane do grupy III metalotionein i są ważnym elementem mechanizmów detoksykacji w obronie roślin przed związkami toksycznymi, szczególnie jonami miedzi oraz kadmu.
18) Fitochrom - fitochrom jest fotoreceptorem, barwnikiem używanym przez rośliny w reakcjach na światło lub jego brak. Maksimum absorpcji barwnika przypada na długości fal odpowiadające światłu czerwonemu i dalekiej czerwieni. Wiele roślin wykorzystuje fitochrom do określenia czasu odpowiedniego na kwitnięcie poprzez określenie dnia i nocy. Zmiany w zawartości poszczególnych form fitochromu pozwalają regulować reakcje związane z cyklem dobowym. Fitochrom reguluje również kiełkowanie nasion, wzrost wydłużeniowy siewek, wielkość i kształt liści, syntezę chlorofilu oraz prostowanie się hipokotyli lub epikotyli siewek roślin dwuliściennych. Cząsteczka fitochromu jest chromoproteiną, składa się z części białkowej połączonej z barwnikiem. Fitochrom zawiera większość roślin, w tym wszystkie rośliny wyższe, a barwniki o bardzo podobnej budowie wykryto także u bakterii. Reakcje roślin będące odpowiedzią na zmiany światła w zakresie spektrum niebieskiego i ultrafioletu regulowane są przez receptory należące do kryptochromów i fototropin.
FORMY FITOCHROMU: Podstawową formą fitochromu jest forma absorbująca światło w zakresie czerwieniu. Maksimum absorpcji przypada na długości fal 650-670 nm. Ta forma fitochromu dla ludzkiego oka posiada barwę turkusowoniebieską. Pochłaniając kwanty światła czerwonego forma PR przekształca się w formę PFR, która wykazuje maksimum absorpcji w tzw. dalekiej czerwieni (długości fal 705-740 nm). Ta forma fitochromu dla oka ludzkiego posiada kolor zielonkawy. Pochłanianie kwantów energii przez formę PFR powoduje jej przekształcenie do formy PR. Fitochrom pochłaniając odpowiedniej długości fale elektromagnetyczne przekształca się w jedną z form, a reakcja jest odwracana przez inną długość fali.

Źródło: Wikipedia. Dwie hipotezy dotyczące przekształceń fitochromu zachodzących podczas absorpcji światła. (PR - Forma absorbująca światło czerwone, PFR - Forma absorbująca w dalekiej czerwieni, B - Białko). Po lewej - odłączenie H+ [1]. Po prawej - tworzenie formy zbliżonej do chlorofilu.
BIOCHEMIA I DZIAŁANIE FITOCHROMU: Chemicznie fitochrom składa się z grupy chromoforowej w postaci pojedynczego, otwartego łańcucha czterech pierścieni pirolowych związanych z białkiem. Grupa chromoforowa pochłaniając światło zmienia konformację chromoproteiny, powodując wzajemne przekształcenie dwóch form fitochromu.
Forma fitochromu PFR zapoczątkowuje w komórce szereg reakcji, między innymi regulując ekspresję niektórych genów. Dokładny mechanizm regulacji metabolizmu komórki przez fitochrom nie został poznany. Forma PR jest wytwarzana w cytozolu i tam pod wpływem światła przekształca się w formę P FR, która przemieszczana jest do jądra komórkowego. Zjawisko to sugeruje bezpośredni udział fitochromu w regulacji ekspresji genów. Istnieje hipoteza, że fitochrom w formie PFR służy jako kinaza wpływająca na aktywację czynników transkrypcyjnych.

HISTORIA BADAŃ NAD FITOCHROMEM: Fitochrom odkryty został przez Sterlinga Hendricksa i Harry'ego Borthwicka z USDA-ARS Beltsville Agricultural Research Center w Marylandzie w latach piećdziesiątych ubiegłego stulecia. Wymienieni naukowcy korzystając z spektrografu zbudowanego z pożyczonych i pozostałych po wojnie części odkryli, że światło czerwone jest bardzo efektywne w pobudzaniu kiełkowania nasion i wywoływaniu kwitnienia roślin. Reakcje na światło czerwone można było odwrócić przez oświetlenie roślin daleką czerwienią. Sam fitochrom został zidentyfikowany za pomocą spektroskopu przez biofizyka Warrena Butlera i biochemika Harolda Siegelmana w roku 1959. To właśnie Butler nadał nazwę fitochrom odkrytej chromoproteinie. W roku 1983 laboratoriach Petera Quaila i Clarka Lagariasa fitochrom został wyizolowany i oczyszczony, a w roku 1985 opublikowali sekwencję genu kodującego fitochrom. W roku 1989 genetycy molekularni wykorzystując przeciwciała monoklonalne wykazali istnienie więcej niż jednej formy fitochromu. W roślinach grochu istnieje forma I obecne w siewkach rosnących w ciemności oraz forma II obecna w zielonych roślinach. Z sekwencjonowania genomu Arabidopsis wiadomo o istnieniu pięciu genów kodujących fitochrom (PHYA - E), u ryżu obecne są jedynie trzy formy (PHYA - C). Istnienie wielu genów kodujących fitochrom w licznych roślinach jedno i dwuliściennychj, prawdopodobnie wiąże się z częstą u roślin poliploidalnością. W związku z tym kukurydza, na przykład, posiada sześć form fitochromu - phyA1, phyA2, phyB1, phyB2, phyC1 i phyC2. Każdy z tych fitochromów posiada inny komponent białkowy, jednak wszystkie posiadają tą samą grupę chromoforową. W późnych latach osiemdziesiątych XX wieku wykazano, że phyA jest degradowany z użyciem ubikwityny. W roku 1996 w zsekwencjonowanym genomie sinicy Synechocystis zauważono istnienie genu o niewielkim podobieństwie do roślinnego fitochromu. Jon Hughes w Berlinie i Clark Lagarias na UC Davis wykazali, że gen ten rzeczywiście koduje barwnik nazwany (Cph1) o właściwościach fitochromu. Przypuszczalnie roślinny fitochrom powstał poprzez przeniesienie genu chloroplastowego do jądra. Fitochrom został także znaleziony w komórkach innych prokariontów np. w Deinococcus radiodurans i Agrobacterium tumefaciens.
U Deinococcus radiodurans fitochrom reguluje wytwarzanie barwników ochronnych, jakkolwiek u Synechocystis i Agrobacterium biologiczna rola tych barwników nie jest znana. W roku 2005 laboratorium Vierstra i Forest Uniwersytetu w Wisconsin opublikowało trójwymiarową strukturę części fotoczułej fitochromu bakterii z rodzaju Deinococcus. Okazało się iż łańcuch białkowy tworzy strukturę o nietypowej formie.
19) Fotoperiodyzm - jest to fizjologiczna reakcja organizmu na zmianę proporcji okresów ciemności i światła w rytmie dobowym związana z działaniem zegara biologicznego. U podstaw fotoperiodyzmu leżą zjawiska fotoindukcji (zaprogramowana genetycznie odpowiedź na skracanie się lub wydłużanie dnia) i fotorefrakcyjności (zachodząca stopniowo adaptacja do skracającego się lub wydłużającego dnia). Okres indukcji świetlnej nazywany jest fotoperiodem, a graniczna wartość czasu trwania dnia lub nocy, uruchamiająca procesy fizjologiczne i behawioralne to fotoperiod krytyczny. Znajomość zjawisk związanych z fotoperiodyzmem jest wykorzystywana w hodowli roślin i zwierząt gospodarskich.
FOTOPERIODYZM U ROŚLIN: Zmiany długości dnia i nocy wpływają na kwitnienie, rozmnażanie wegetatywne, tworzenie organów spichrzowych oraz przechodzenie roślin w stan spoczynku. Najlepiej poznanym zjawiskiem związanym z fotoperiodyzmem jest kwitnienie. Zjawisko indukcji kwitnienia przez zmiany w długości dnia i nocy zaobserwowali Gardner i Allard, dwaj amerykańscy naukowcy, którzy w roku 1920 otrzymali w wyniku mutacji odmianę tytoniu o nazwie Maryland Mammoth. Roślina rozwijała się do wysokości 3m, jednak nie zakwitała przez cały sezon wegetacyjny. Jedynie po przeniesieniu do szklarni, zakwitała w okresie Bożego Narodzenia. Dalsze badania udowodniły, że kwitnienie może być indukowane odpowiednią długością dnia lub nocy.
W zależności od rodzaju reakcji wyróżnia się rośliny:
- długiego dnia - kwitnące, gdy okres światła (długość dnia) przekroczy wartość krytyczną, specyficzną dla poszczególnych gatunków (np. szpinak, zboże, trawy, hortensja, rzodkiewka, burak pastewny i cukrowy, marchew),
- krótkiego dnia - kwitnące, gdy okres światła (długość dnia) będzie krótszy niż wartość krytyczna (np. tytoń, kukurydza, proso, ryż, astry ,chryzantemy, rzepak, wilczomlecz, soja),
- obojętne - kwitnące niezależnie od długości dnia i nocy (np. gryka, pomidor, ogórek, groch, bób, ryż - odmiany jare).
Przerwanie okresu ciemności zapobiega kwitnieniu roślin dnia krótkiego, a indukuje kwitnienie u roślin dnia długiego. Kluczowa jest zatem długość okresu ciemności. Fotoreceptorem biorącym udział w odbieraniu informacji o długości dnia i nocy jest fiochrom, który przechodzi w ciemności z formy PFR do formy PF. Na świetle zachodzi proces odwrotny. U roślin dnia krótkiego do zakwitnięcia konieczny jest odpowiednio niski poziom PFR, a u roślin dnia długiego poziom PFR musi być odpowiednio wysoki. Fitochrom może bezpośrednio wpływać na ekspresję genów związanych z kwitnieniem, rozważano także istnienie hormonu - florigenu, który byłby bezpośrednim induktorem kwitnienia. Fotoperiodyzm ma także wpływ na wykształcanie bulw, rozłogów, opadanie liści, mrozoodporność roślin, przechodzenie w stan zimowy drzew oraz krzewów.
 |
| Źródło: Wikipedia. Fotoperiodyzm u roślin - tytoń - Nicotiana tabacum, jest rośliną dnia krótkiego, przy długim dniu rozwija się tylko wegetatywnie. |
 |
| Źródło: Wikipedia. Beta-Sitosterol. |
20) Fitosterole, sterole roślinne - jest to grupa steroli wytwarzanych przez rośliny. Znanych jest ponad 150 różnych steroli roślinnych. Sterole wytwarzane przez rośliny są elementem strukturalnym błon komórkowych i cytoplazmatycznych, uczestniczą w regulacji procesów wewnątrzkomórkowych oraz pełnią funkcję substancji obronnych przed roślinożercami. Wiele steroli roślinnych po połączeniu z cukrem tworzy glikozydy, będące substancją obronną oraz znajdujące zastosowanie jako leki.


FUNKCJE: Wolne i pochodne steroli roślinnych obecne są w błonach komórkowych i cytoplazmatycznych. Zawartość poszczególnych fitosteroli jest różna dla błon różnych organelli. Błona komórkowa zawiera głównie sitosterol, kampesterol i stigmasterol. W błonach jądrowych występuje cholesterol. Sterole z grupy ekdysonów są hormonami wywołującymi wylinkę owadów. Obecne w roślinach fitosterole mogą zaburzać cykl rozwojowy owadów. Związki z grupy limonoidów są bardzo skutecznym środkiem odstraszającym owady. Glikozydy steroli oraz ich pochodne tworzą grupę związków nazywanych saponinami wykazującymi właściwości detergentów. Zdolność do rozbijania błon skutkuje wysoką toksycznością dla roślinożerców. Właściwość ta została także wykorzystana przez człowieka przy produkcji środków piorących z surowców roślinnych. Motyle z rodzaju Danaus wykształciły odporność na glikozydy obecne w roślinach, w efekcie gąsienice gromadząc w organizmie toksyczne związki pochodzące ze zjadanych liści, stają się trujące dla ptaków.
 |
| Źródło: Wikipedia. Nasiona pomidora wykazują fotoblastię ujemną, czyli kiełkują gdy dostęp do światła jest odcięty. |
21) Fotoblastia, nasiona fotoblastyczne - jest to zjawisko regulacji kiełkowania nasion przez światło występujące u niektórych roślin wyższych odpowiedzialne za pozostanie w stanie spoczynku lub wyjście z niego. W zależności od reakcji na światło rozróżniana jest fotoblastia dodatnia, gdy światło stymuluje kiełkowanie i fotoblastia ujemna, gdy światło hamuje kiełkowanie. Tylko nieliczne rośliny wytwarzają nasiona niewrażliwe na światło. Jest to około 4% gatunków, jednak jest wśród nich wiele gatunków uprawianych przez człowieka. Światło może być czynnikiem niezbędnym do wykiełkowania, tak jest u jemioły lub jedynie przyspieszać kiełkowanie.
MECHANIZM: Zjawisko fotoblastii zostało najlepiej poznane u sałaty (Lactuca sativa). Nasiona tej rośliny kiełkują po ekspozycji na światło. Za odbiór bodźca odpowiedzialny jest fitochrom. Indukcja kwitnienia następuje na skutek oświetlenia światłem czerwonym. Daleka czerwień może odwracać efekt indukcji. Odbiór bodźca przez fitochrom prowadzi do zmian w poziomie giberelin (GA) i kwasu abscysynowego (ABA). Podstawowym czynnikiem decydującym o wyjściu ze stanu spoczynku jest synteza giberelin. Do pobudzenia kiełkowania wystarczy natężenie 10 J m-2 co odpowiada kilkuminutowemu oświetlaniu światłem księżyca lub błyskowi światła słonecznego trwającemu 0,1 s.
22) Fototropiny - roślinne receptory światła niebieskiego. Razem z kryptochromami i fitochromami pozwalają roślinom na reakcje na zmieniające się warunki oświetleniowe. Fototropiny to flawoproteiny (flawoproteina to białko zawierające pochodną ryboflawiny. Flawoproteiny biorą udział w katabolicznych procesach oksydacyjnych. Najczęstszą formą koenzymu flawoprotein jest FAD), które absorbują światło niebieskie poprzez związany niekowalencyjnie barwnik - FMN (Mononukleotyd flawinowy - organiczny związek chemiczny, ester kwasu fosforowego i rybloflawiny).

Fototropiny zawdziędzają swoją nazwę roli jaką odgrywają w kontrolowaniu fototropizmu czyli wyginania się roślin w stronę światła. Oprócz tej reakcji, fototropiny powodują otwieranie aparatów szparkowych, są odpowiedzialne za wywoływanie światłem ruchu chloroplastów, kontrolują także rozwój blaszki liściowej. Roślina modelowa rzodkiewnik pospolity (Arabidopsis thaliana) ma dwa geny kodujące fototropiny: 1 i 2. Inne gatunki roślin mogą mieć ichwięcej. Obie fototropiny różnią się wrażliwością na światło - fototropina 1 jest bardziej czuła - wywołuje jednak podobne reakcje w różnym zakresie natężeń światła. Wyjątkiem jest reakcja ucieczki chloroplastów, którą kontroluje wyłącznie fototropina 2. Fototropina 1 reguluje natomiast wywołane światłem hamowanie wzrostu hipokotyla. Fotropiny należą do kinaz białkowych aktywowanych światłem niebieskim. Światło powoduje fosforylację fototropin obecnych w błonie komórkowej, co prowadzi do zapoczątkowania łańcucha przekazywania sygnału wewnątrz komórki.

WIĘCEJ INFORMACJI O FOTOTROPINACH: TUTAJ.
23) Hartowanie roślin - Hartowanie roślin to zjawisko sezonowej aklimatyzacji roślin do niskich temperatur. Proces tne jest formą odporności na niskie temperatury i prowadzi do stopniowego nabycia mrozoodporności. Hartowanie zachodzi w dwóch lub trzech etapach.
ETAP PIERWSZY: Etap pierwszy rozpoczyna się w wyniku odebrania przez roślinę informacji o skracaniu dnia. Za informacje o fotoperiodzie odpowiedzialne są przemiany fitochromu. Drugim odbieranym bodźcem jest zmiana temperatury. U drzew hartowanie rozpoczyna się w temperaturze 5-10 stopni Celsjusza, a u roślin dwuletnich 2-5 stopni Celsjusza. Po odebraniu sygnału w roślinie wzrasta stężenie kwasu abscysynowego i spada zawartość giberelin. W komórkach parenchymatycznych gromadzone są substancje zapasowe, głównie skrobia i lipidy. W żywych komórkach redukcji ulega objętość wakuoli. Maleje też potencjał wody w komórce. U roślin dwuletnich dochodzi do zmian morfologicznych pojawiających się w efekcie względnego nadmiaru energii świetlnej pochłanianej przez chlorofil, jednakże nadmiar ten nie może być wykorzystany do syntezy związków organicznych w cyklu Calvina z powodu spadku aktywności enzymów w niskich temperaturach. Powstają liście o grubszych blaszkach i zbudowane z komórek o zwiększonej grubości ścian komórkowych.
ETAP DRUGI: Drugi etap hartowania indukowany jest spadkiem temperatur poniżej 0 stopni Celsjusza. Podczas tego etapu następuje przebudowa błon cytoplazmatycznych. Kwasy tłuszczowe nasycone wymieniane są na kwasy tłuszczowe nienasycone. Zmienia się proporcja fosfatydylocholiny, fosfatydylogliceroli i fosfatydyloinozytolu. W komórkach wytwarzane są białka indukowane chłodem oraz aktywowane enzymy antyoksydacyjne. W tkankach wzrasta stężenie etylenu. Modyfikacji ulega metabolizm, rośliny dwuletnie przechodzą w stan spoczynku wymuszonego, a drzewa i byliny w stan spoczynku głębokiego.
ETAP TRZECI: W trzecim, w warunkach mrozu, dochodzi do krystalizacji wody w przestrzeniach międzykomórkowych oraz naczyniach. Dochodzi także do ubytku wody w cytoplazmie. Rośliny są w stanie przetrwać mrozy do -25 stopni Celsjusza, a drzewa nawet do -50 stopni Celsjusza. Wiosną, gdy temperatury znów osiągają wartości dodatnie dochodzi do rozhartowania roślin. Rozhartowane rośliny są wrażliwe nawet na przymrozki. Do rozhartowania roślin drzewiastych może dojść także w środku zimy, jeśli nastąpi wyraźne ocieplenie. Im dłuższy jest okres ocieplenia i wyższa temperatura dobowa tym proces rozhartowania może być głębszy, łącznie z cofnięciem zmian zachodzących w drugiej fazie hartowania. Jeżeli po okresie ocieplenia spadki temperatur są stopniowe to rośliny ponownie nabywają wyższą odporność na mróz.
24) Imbibicja - jest to proces pęcznienia koloidów. Polega on na przyłączaniu coraz większej ilości cząsteczek wody wiązaniami wodorowymi do cząsteczek płaszcza wodnego otaczającego cząsteczkę koloidu hydrofilnego, a tym samym na zwiększaniu odległości między kolejnymi micellami koloidowymi. Występuje m.in. w początkowej fazie kiełkowania nasion - pęcznieniu, w której zwiększają swoją objętość i masę po zanurzeniu w wodzie lub otoczeniu wilgotnego środowiska. Zjawisko to zachodzi również w stomatologicznych masach wyciskowych, przechowywanych w środowisku hipotonicznym (np. w wodzie).


Źródło: uwm.edu.pl
25) Immunia, odpornośc całkowita - jest to zjawisko całkowitej odporności roślin na biotyczne czynniki stresowe. Mechanizmy odporności całkowitej powodują, że patogeny nie są w stanie wniknąć do organizmu rośliny, a roślinożercy mają całkowicie uniemożliwione żerowanie. Immunia jest zjawiskiem częstym u dziki rosnących roślin, stąd rzadko występują u takich roślin objawy chorób. Wrażliwe na atak patogenu mogą być rośliny w określonej fazie rozwojowej.
26) Inkrustacja - jest to proces odkładania się niektórych substancji w szkielecie celulozowym pierwotnej ściany komórkowej roślin. Do substancji tych należy m.in. lignina (drewnienie), garbniki, żywice, olejki eteryczne, białka i niektóre związki mineralne, jak węglan wapnia, krzemionka (mineralizacja). Obecność ligniny w ścianie powoduje jej silne stwardnienie i znaczne zmniejszenie zawartości wody. Ściana staje się słabo przepuszczalna dla wody i powietrza (co często jest przyczyną śmierci protoplastu), równocześnie roślina nabiera sztywności i mechanicznej odporności.
27) Inteligencja roślin - są to złożone zachowania adaptacyjne występujące u roślin. Zachowania te umożliwiają organizmom roślinnym dostosowanie się do warunków środowiska. Mechanizmy adaptacyjne, będące plastyczną reakcją fenotypową w odpowiedzi na sygnały ze środowiska powstały w wyniku ewolucji i obejmują zachowania takie jak reakcja na stres lub obrona przed patogenem. Właściwe reagowanie na zmieniające się warunki wymaga istnienia pamięci, uczenia się i inteligencji. Reakcje roślin mogą mieć charakter morfologiczny lub fizjologiczny. Koncepcja zachowań inteligentnych u roślin została zapoczątkowana przez Karola Darwina.
HISTORIA: Możliwość istnienia życia wewnętrznego u roślin rozważał Arystoteles. Linneusz postulował istnienie wzrostu u kamieni, wzrostu i życia u roślin oraz życia i czucia u zwierząt. Istnienie centrum inteligencji u roślin odpowiadającego mózgowi zwierząt zostało zaproponowane przez Darwina. Centrum takie miało być zlokalizowane w wierzchołku korzenia i odpowiadać za przetwarzanie sygnałów odbieranych przez komórki czuciowe. Zdolność do odbierania sygnałów przez organizm roślinny została następnie opisana w podręczniku Rabera "Principles of Plant Physiology" w roku 1933. W drugiej połowie XX wieku znane już były reakcje roślin na chemiczne i fizyczne sygnały ze środowiska i zaobserwowano także powstawanie potencjałów czynnościowych u łubinu. Przenoszenie informacji w postaci zmian potencjału umożliwiają wiązki przewodzące. Sygnały elektryczne indykowane są w wyniku zranienia oraz w warunkach gwałtownego stresu. Doświadczenia potwierdziły wpływ sygnałów elektrycznych na intensywność fotosyntezy. Znanych jest szereg endogennych substancji chemicznych pełniących rolę regulatorową. Część z nich to fitohormony, których rola w regulacji reakcji morfologicznych i fizjologicznych jest stosunkowo dobrze znana. W roślinach wykryto także wiele substancji, które u zwierząt pełnią funkcję neurotransmiterów. Funkcja tych związków w komórkach roślinnych jest słabo poznana. Związkiem zaangażowanym w wiele reakcji organizmów roślinnych jest tlenek azotu (NO) uczestniczący w regulacji wzrostu korzeni bocznych, geotropizmie, przerywaniu spoczynku nasion, kiełkowaniu, produkcji fitoaleksyn, hamowaniu starzenia i wielu innych.
PRZEJAWY INTELIGENCJI ROŚLIN: Tradycyjne rozumienie inteligencji związane jest z obserwowaniem ruchu, jednak każde zachowanie o charakterze adaptacji może być traktowane jako przejaw inteligencji. Główną różnicą w badaniach nad reakcjami zwierząt i roślin jest skala czasowa, która u roślin obejmuje tygodnie lub miesiące. Podstawowym zachowaniem inteligentnym jest uczenie się. Wymaga ono osiągania celu przy jednoczesnej korekcji błędów. Rośliny muszą wykazywać właściwą reakcję na siłę grawitacji, natężenie światła, stężenie soli mineralnych i zawartość wody w środowisku. W przypadku grawitropizmu obserwowano odchylenie rosnących organów, które początkowo przekraczało potrzebę. Kolejne oscylacyjne ruchy wzrostowe umożliwiały dopasowanie reakcji do potrzeb. Zachodziła korekcja błędów w stosunku do celu. Przejawy uczenia się były obserwowane także w ruchach aparatów szparkowych, które muszą zapewnić dostosowanie do ilości wody dostarczanej przez korzenie.
MECHANIZMY INTELIGENTNYCH REAKCJI NA BODŹCE: Uczenie się wymaga istnienia systemu odbioru i przekazywania sygnałów. Badania koncentrują się na poszukiwaniu receptorów i cząstek sygnałowych. Na podstawie analizy genomu Arabidopsis oszacowano, że w roślinie istnieje prawie tysiąc kinaz białkowych. Potwierdzono także występowanie w komórkach roślinnych szeregu cząstek uczestniczących w transdukcji sygnału.
27) Jednostkowa produktywność liści:
 |
| Źródło: Wikipedia. |
28) Kairomon (z gr. kairos - korzyści) - jest to substancja semiochemiczna wydzielana mimowolnie przez organizm niosąca sygnał neutralny lub niekorzystny dla nadawcy, a korzystny dla odbiorcy. Przykłądowo - jako kairomon może działać zapach drapieżnika wyczuwany przez jego ofiarę. Ofiara po detekcji takiej substancji może podjąć mechanizmy obronne. Innym przykładem kairomonów są substancje wydzielane przez rośliny, które przyciągają roślinożerców. Zdarza się, że dana substancja jest kairomonem w stosunku do jednego gatunku (jako jego atraktant), a allomonem dla innego (jako jego repelent). Istnienie kairomonów udowodniono w przypadku ryb i zooplanktonu (zapach ryb drapieżnych indukuje mechanizmy obronne drobnych ryb, zapach ryb planktonożernych i bezkręgowców drapieżnych indukuje mechanizmy obronne - morfologiczne (cyklomorfoza) i behawioralne - u zwierząt planktonowych). Podobnie jest przy wykrywaniu kairomonów pochodzących od zooplanktonu przez organizmy fitoplanktonowe.
29) Koliny - są to substancje chemiczne wytwarzane przez rośliny wyższe wpływające na wzrost i rozwój roślin wyższych rosnących w pobliżu. Jest to grupa związków zaliczanych do allelopatin i w znacznym stopniu decydują one o składzie gatunkowym formacji roślinnych. Substancje allelopatyczne mogą być wytwarzane przez tkankę gruczołową organów nadziemnych i uwalniane w formie lotnej lub rzadziej w wyniku eksudacji przez system korzeniowy.
 |
| Źródło: ks.uiuc.edu - Struktura Kryptochromu. |
HISTORIA ODKRYCIA: Właściwości molekularne kryptochromu opisano po raz pierwszy pod koniec 1993 roku. Przedmiotem badań był mutant hy4 rzodkiewnika pospolitego (Arabidopsis thaliana), posiadający zaburzenia w postaci braku hamującego wpływu światła niebieskiego na wzrost hipokotylu.
BUDOWA I FUNKCJA: Kryptochromy są obecne u wszystkich gatunków roślin. Jest to polipeptyd, który u Arabidopsis thaliana ma masę cząsteczkową równą 75,8 kDa i składa się z 681 aminokwasów. N-końcowy fragment tego polipeptydu wykazuje dużą homologię z bakteryjnymi fotoliazami, natomiast C-końcowy fragment ma budowę zbliżoną do tropomiozyny A z mięśni gładkich szczura. Część środkowa nie wykazuje żadnego podobieństwa do znanych dotąd polipeptydów. U większości roślin występują dwa homologiczne kryptochromy: Cry1 i Cry2. Kryptochromy Cry rzodkiewnika, posiadają dwa chromofory: pterynę i flawinę (substancja zbliżona do pteryny). Chromofor pterynowy jest tym, który bezpośrednio absorbuje foton, co następnie powoduje emisję przez niego energii, która z kolei jest absorbowana przez flawinę. Ta prawdopodobnie pośredniczy w fosforylacji określonej domeny w kryptochromie, tym samym rozpoczynając kaskadę transdukcji sygnału, finalnie wpływającego na regulację ekspresji genów w jądrze komórkowym. Produkty genów CRY1 i CRY2 uczestniczą w procesie deetiolacji. Poziom Cry2 po naświetleniu światłem niebieskim, ultrafioletowym (UV-A:320-390 nm) i zielonym szybko się obniża. W warunkach małego natężenia oświetlenia u rzodkiewnika deetiolację kontrolują obydwa typy kryptochromów, natomiast w środowisku o dużym natężeniu oświetlenia, główną funkcję w regulacji tego procesu pełni Cry1.
 |
| Źródło: riken.jp - Funkcje pełnione przez Kryptochrom. |
WIĘCEJ INFORMACJI NA TEMAT KRYPTOCHROMU ZNAJDZIESZ: TUTAJ.
31) Ksenia (z gr. xenos - obcy, gość) - jest to zjawisko ujawniania się genotypu rośliny ojcowskiej w tkankach nasion lub owoców. Występowanie cech będących wynikiem ekspresji genów zawartych w pyłku w tkankach, które pochodzą wyłącznie z rośliny matczynej określane jako metaksenia, zaś w tkankach, które powstają z połączenia gamet - gamoksenia. Termin został wprowadzony przez Wilhelma Olbersa Focke w roku 1881. Początkowo rozważano także wpływ zarodka na bielmo, które uznawane było za tkankę pochodzącą z rośliny matczynej. Po odkryciu podwójnego zapłodnienia u okrytonasiennych, stało się jasne, że w tych komórkach obecne są geny pochodzące z pyłku. Poznanie podwójnego zapłodnienia nie pozwoliło jednak wyjaśnić wszystkich obserwowanych zjawisk związanych z ksenią. Wpływ zarodka na właściwości owoców jest prawdopodobnie związany z wydzielanymi hormonami, szczególnie auksyną Dla pierwszych naukowców opisujących zjawisko techniki pozwalające potwierdzić taką hipotezę nie były dostępne. Zjawisko znajduje zastosowanie w rolnictwie i ogrodnictwie, gdzie dobór rośliny z której pochodzi pyłek może zapewniać korzystny dla człowieka kształt i kolor owoców, wielkość komórek nasion, czas rozwoju i skład chemiczny oraz wpływać na ilość plonu.
EFEKT W NASIONACH: Ksenia może być wykorzystana do zwiększenia masy nasion oraz rozmiarów zarodka. W przypadku bawełny zjawisko może mieć korzystne efekty dla materiału siewnego oraz umożliwiać zwiększenie ilości oleju uzyskiwanego z nasion. W badaniach wykazano także wpływ ksenii na zawartość i skład oleju różnych odmian migdałów.
WYKORZYSTANIE W ROŚLINACH TRANSGENICZNYCH: Ze względu na pojawiające się w społeczeństwie obawy związane z konsekwencjami rozsiewania pyłku roślin modyfikowanych genetycznie rozważana jest możliwość uprawy modyfikowanych roślin męskosterylnych i wykorzystywanie do zapylania roślin niemodyfikowanych. Zjawisko ksenii może dać dodatkowe korzyści z takiego rozwiązania, umożliwiając zwiększenie plonów.
32) Kutykularyzacja - jest to jeden z typów adkrustacji prowadzący do tworzenia na powierzchni nadziemnych organów roślinnych kutykuli. Proces polega na odkładaniu kutyny na zewnętrznej powierzchni ściany komórkowej komórek tworzących skórkę pędu. Następuje w wyniku wydzielania poprzez ściany drobnocząsteczkowego prekursora kutyny, który na powierzchni ulega polimeryzacji prowadzącej do wytworzenia ciągłej warstwy ochronnej. W odróżnieniu od kutynizacji proces tworzenia kutyny następuje na zewnątrz ścian komórkowych.
33) Kutynizacja - proces polegający na odkładaniu kutyny wewnątrz ściany komórkowej, między warstewkami celulozy. W jego wyniku powstaje warstwa kutykularna (skutykularyzowana) ściany komórkowej. Proces ma charakter pośredni między adkrustacją i inkrustacją. Podobny proces kutykularyzacji polega na wydzielaniu kutyny poza ścianę komórkową komórek zewnętrznych rośliny - prowadzi do wytworzenia warstwy ograniczającej parowanie rośliny - kutykuli.
34) Maceracja.
 |
| Źródło: Wikipedia. |
35) Marazminy - są to substancje chemiczne wytwarzane przez mikroorganizmy wpływające na wzrost i rozwój roślin wyższych rosnących w pobliżu. Jest to grupa związków zaliczanych do allelopatin.
36) Materiał zapasowy roślin - są to związki organiczne gromadzone przez roślinę i zużywane na niektórych etapach rozwoju, np. w czasie kiełkowania nasion lub w okresach niekorzystnych dla rośliny warunków środowiska. Związki organiczne wytworzone w procesie fotosyntezy tylko w pewnym stopniu są zużywane podczas procesów metabolicznych, część z nich jest odkładana u wykorzystywana w odpowiednim momencie. W niektórych przypadkach duże ilości substancji zapasowych mogą być magazynowane w organach spichrzowych rośliny.
Materiały zapasowe gromadzone są w cytozolu i organellach komórkowych. Każda żywa komórka ma zdolność do gromadzenia substancji zapasowych. Najczęściej funkcję tkanki spichrzowej spełniają komórki miękiszowe. Materiały zapasowe stanowić mogą węglowodany (glukoza, sacharoza, skrobia, inulina, hemiceluloza), białka zapasowe (np. w postaci ziaren aleuronnowych), tłuszcze. Najczęściej materiałem zapasowym są węglowodany.
 |
| Źródło: Wikipedia. Ziarna skrobi w komórkach bulwy ziemniaka. |
 |
| Źródło: img.guidechem.com - Chloroflurenol to przykład morfaktyny. |
 |
| Źródło: Wikipedia. Cząsteczka Fluorenu. |
38) Myrmekotrofia - jest to forma mutualizmu, polegająca na pozyskiwaniu przez rośliny związków mineralnych z odpadów kolonii mrówek z tymi roślinami związanych, takich jak odchody czy szczątki owadów. Martwe szczątki stanowią dodatkowe źródło azotu. Dojrzałe osobniki Nepenthes bicalcarata mogą uzyskiwać około 40% azotu występującego w liściach ze współpracy z mrówkami. Rośliny wykształcają specjalne struktury ułatwiające osiedlenie się mrówek. Są to modyfikacje liści, ogonków liściowych, łodyg oraz puste ciernie. Żyjące w takich strukturach mrówki nie tylko przyczyniają się do zwiększenia dostępności związków azotowych, lecz również zapewniają roślinom obronę przed owadami zjadającymi liście. Rośliny pozyskujące azot w wyniku myrmekotrofii występują zwykle w ekosystemach gdzie dostępność tego pierwiastka jest ograniczona. Tę strategię ewolucyjną stosują m.in. epifityczne rośliny z rodzaju Myrmecodia.
Źródło: Wikimedia.
39) Nektar - jest to wydzielina miodników (nektarników) roślin. Jest to wodny roztwór cukrów, głównie fruktozy i glukozy. Nektar wabi zwierzęta zapylające kwiaty (głównie owady), przez co odgrywa kluczową rolę w procesie rozmnażania roślin. Jego skład i intensywność wydzielania są zmienne u poszczególnych gatunków, o różnej porze dnia i w zależności od warunków pogodowych. Z nektaru pszczoły wytwarzają miód, który powstaje w wyniku zagęszczenia nektaru (odparowania znacznej części wody), rozłożenia scharozy na cukry proste i zakonserwowania powstałej substancji niewielką ilością kwasu mrówkowego.
SKŁAD CHEMICZNY I WŁAŚCIWOŚCI FIZYCZNE: Skład i własności nektaru są bardzo zmienne. Stałym składnikiem są węglowodany - glukoza, fruktoza i sacharoza. Ten ostatni dominuje w nektarze kasztanowców i robinii, podczas gdy na przykład w nektarze gryki występują wyłącznie cukry proste. Zawartość cukrów waha się u różnych gatunków od ok. 5%-8% u szachownicy cesarskiej, do ok. 70% u kasztanowca i 76% u lebiodki pospolitej. Od zawartości cukrów zależna jest gęstość nektaru. Nektar zawiera poza tym niewielkie ilości soli mineralnych (zwłaszcza potasu), olejki eteryczne, kwasy, zasady, barwniki oraz aminokwasy. W większości przypadków brak jest w nektarze substancji zapachowych (do wyjątków należy np. roślina Nicotiana attenuata, która zawiera benzyloaceton - silną substancję zapachową).
W skład nektaru wchodzić mogą w przypadku niektórych gatunków roślin składniki trujące dla pszczół lub ludzi. Nektar bielunia indiańskiego i stworzony z niego miód zawierają skopolaminę. Trujący jest również nektar różanecznika żółtego i tojadów. Część składników zawarta w nektarze, zarówno szkodliwych jak i leczniczych, zanika w procesie dojrzewania miodu.
POWSTAWANIE: Nektar wytwarzany jest przez rośliny w gruczołach zwanych miodnikami lub nektarnikami. Płyn wycieka na zewnątrz miodników albo w wyniku dyfuzji przez ścianę komórkową. Gdy miodniki są okryte skórką, nektar wycieka przez szparki, kanaliki lub pęknięcia skórki. Różna jest wydajność tworzenia nektaru przez różne rośliny. Jeden kwiat trojeści amerykańskiej wytwarza go dziennie w ilości 5,3 mg, lipy szerokolistnej - 2,3 mg. Młode kwiaty zwykle nie mają nektaru, wytwarzany jest w większej ilości w miarę trwania kwitnienia. Wydajność procesu tworzenia tej wydzieliny przez rośliny zależną jest od pory dnia (zwykle największa jest w godzinach popołudniowych) i temperatury (większość roślin do wytworzenia nektaru wymaga temperatur powyżej 10-12 stopni Celsjusza). Podczas suszy zwykle spada wydajność wytwarzania nektaru.
ZNACZENIE: Nektar odgrywa kluczową rolę w tworzeniu symbiozy między różnymi grupami zwierząt dokonującymi zapylenia a roślinami kwiatowymi. Owady, ale także ptaki, nietoperze i niektóre torbacze, korzystają z niego jako pożywienia, w wielu wypadkach specjalizując się i uzależniając od niego. W zamian przy okazji odwiedzin kwiatów dokonują przenosin między nimi pyłku, dzięki czemu mimowolnie je zapylają. Owadami wyspecjalizowanymi w zbiorze nektaru są m.in. pszczoły. Gromadzą one i przechowują nektar poddając go obróbce za pomocą swojej śliny, tworząc miód w wyniku procesu dojrzewania takiej mieszaniny. Stanowi on wysokoenergetyczne pożywienie, podbierane pszczołom także przez inne zwierzęta. Miód jest pożywieniem cenionym także przez ludzi. W celu zwiększenia jego dostępności prowadzona jest działalność pszczelarska. W jej ramach m.in. rozpowszechniane są rośliny miododajne. Znajdujące się wśród nich rośliny nektarodajne wyróżniają się dużą wydajnością wytwarzania nektaru oraz dużą zawartością w nich cukrów.
40) Obrona przed roślinożercami -jest to zespół mechanizmów w postaci przystosować morfologicznych oraz fizjologicznych roślin, mających na celu przeciwdziałanie lub zniwelowanie skutków ataku organizmów roślinożernych. Mechanizmy te mogą mieć charakter bezpośredniego oddziaływania między rośliną a żywiącym się nią organizmem lub charakter pośredni polegający na współpracy z naturalnymi wrogami roślinożerców. Powstanie mechanizmów obronnych w wyniku ewolucji miało miejsce po pojawieniu się owadów, które wykorzystywały rośliny jako źródło pokarmu. Koewolucja roślin i owadów zachodzi od permu, jednak gwałtowny wzrost wzajemnych adaptacji obejmuje okres kredy, kiedy to pojawiły się rośliny okrytonasienne.
OBRONA BEZPOŚREDNIA: Obronę mechaniczną (fizyczną), bezpośrednią zapewniają struktury wytwarzane przez wiele gatunków roślin takie jak włoski, trichomy, ciernie, kolce i zgrubiałe liście. Zniechęca roślinożerców również silne owłosienie. Przed żerowaniem ślimakó∑ bronią rośliny rafidy - igiełkowate kryształy szczawianu wapnia. Niektóre gatunki unikają w końcu zgryzania ściśle przylegając do podłoża. Mechanizmy fizjologiczne (chemiczne) polegają głównie na syntezie metabolitów wtórnych oraz białek toksycznych lub odstraszających. Terpenoidy, alkaloidy, antocyjany, związki fenolowe oraz chinony mogą powodować śmierć zwierząt, negatywnie wpływać na ich rozwój, odstraszać lub zaburzać metabolizm. Mimo początkowych wątpliwości, czy metabolity wtórne są produktem wytwarzanym specjalnie w celu obrony przed roślinożercami, czy są tylko wykorzystywanym tak produktem ubocznym metabolizmu, przeważa ten pierwszy pogląd. Chemiczna obrona może mieć charakter konstytutywny (związki szkodliwe dla roślinożerców wytwarzane są przez roślinę przez cały czas) lub być indukowana przez atak. W drugim przypadku szkodliwe związki chemiczne wytwarzane są w efekcie uszkodzenia rośliny. Za indukcje reakcji obronnej roślin mogą odpowiadać elicytory obecne już w jajach składanych przez owady.
- *Elicytory - to związki chemiczne należące do oligosacharydów, glikopeptydów i glikoprotein, białek, lipidów, które indukują biochemiczne reakcje obronne roślin. Wydzielane są przez ściany komó®kowe zaatakowanej rośliny lub przez mikroorganizm patogeniczny. Elicytory biorą również udział w reakcjach obronnych roślin skierowanych przeciw roślinożercom. Efektem pojawiania się elicytorów w organizmie roślinnym są reakcje morfologiczne i fizjologiczne umożliwiające jej przetrwanie i konkurencyjność. Związki pojawiające się w wyniku uszkodzeń mechanicznych powodowanych przez roślinożerne owady u niektórych roślin stymulują wytwarzanie nikotyny, będącej silną toksyną dla tych zwierząt.
OBRONA POŚREDNIA: W przypadku obrony przed żywiącymi się roślinami owadami, organizmy żywicielskie wytwarzają substancje chemiczne przywabiające drapieżniki zjadające roślinożerców. Pośrednia ochrona przed żywiącymi się roślinami owadami jest coraz częściej badana. Ma to związek z poszukiwaniem przyjaznych dla środowiska metod ochrony roślin wykorzystywanych w rolnictwie. Wiele roślin skutecznie broni się przed owadami roślinożernymi poprzez współpracę z organizmami drapieżnymi lub pasożytniczymi, pełniącymi rolę agentów chroniących przed roślinożercami. Rośliny wabią mutualistów za pomocą lotnych związków organicznych albo wydzielanie nektaru. Lotne związki organiczne służą do wabienia głównie drapieżnych roztoczy i pasożytniczych os. Wydzielany nektar przyciąga przede wszystkim mrówki. Wytworzenie substancji wabiących następuje w efekcie działania hormonu roślinnego, kwasu jasmonowego w reakcji na uszkodzenia powstające podczas żerowania owadów roślinożernych. Wykazano znaczący udział obrony z udziałem mrówek roślin trawiastych ekosystemów Brazylii. Mrówki zapewniają skuteczną obronę przed gąsienicami. Motyle unikają składania jaj na roślinach bronionych przez mrówki.
Brak komentarzy:
Prześlij komentarz