
 |
| Źródło: Wikipedia. Apokarpiczne słupko- -wie u jaskra rozłogowego. |
1) Apokarpie - słupkowie apokarpiczne, zalążnia apokarpiczna - to rodzaj słupkowia u roślin, u których tworzone jest ono z więcej aniżeli jednego owocolistka, przy czym każdy z owocolistków pozostaje wolny (nie zrośnięty z innymi). W trakcie rozwoju (ontogenezy) poszczególne owocolistki zwijają się i zrastają wzdłuż swych brzegów (tworząc szew brzuszny). W efekcie w kwiecie powstaje wiele jednoowocolistkowych słupków, tym samym także wiele zalążni. Po zapyleniu i zapłodnieniu zalążków z zalążni apokarpicznych rozwijają się owoce zbiorowe.
2) Dno kwiatowe/ oś kwiatowa (łac. receptaculum) - jest to element budowy kwiatu roślin okrytonasiennych będący przeważnie mniej lub bardziej rozszerzonym końcem szypułki. Na dnie kwiatowym osadzone są wyrastające spiralnie lub w okółkach pozostałe elementy budowy kwiatu - okwiat, pręcikowie i słupkowie. Dno kwiatowe stanowi twór homologiczny do osi kłosa zarodnionośnego roślin zarodnikowych i osi strobili nagonasiennych.
 |
| Źródło: Wikipedia. Różne ukształtowanie dna kwiatowego i osadzenie słupka. 1 - słupek górny, 2 - słupek pośredni, 3 - słupek dolny.
W zależności od ukształtowania dna kwiatowego i osadzenia na nim słupkowia wyróżnia się:
Silnie wgłębione, często mięsiste dno kwiatowe, otaczające słupki znajdujące się wewnątrz niego określane jest mianem hypancjum - jest to element budowy niektórych kwiatów; dno kwiatowe o specyficznej budowie - silnie wgłębione lub posiadające wąską gardziel. Wskutek takowej budowy słupki znajdują się wewnątrz dna kwiatowego, są ukryte i pozornie wydaje się, że są dolne. Tak jest np. u róży. Czasami, jak np. u wiesiołka dno kwiatowe wyrasta w górnej części zalążni, tak że szyjka słupka znajduje się wewnątrz niego. Z kwiatów posiadających hypancjum często powstaje owoc pozorny.

Źródło: Wikipedia. Kwiat róży - wewnątrz hypancjum oznaczonego ciemnozielonym kolorem znajdują się słupki oznaczone różowym kolorem.
Mięsiste dno kwiatowe u wielu roślin bierze udział w tworzeniu owoców, zwanych w takim przypadku owocami pozornymi. Wypukłe dno kwiatowe, w którym osadzone są drboen orzeszki tworzy np. owoc pozorny truskawki. Wypukłe dno otoczone pestkowcami wchodzi w skłąd owocu pozornego malin. Zmięśniałe hypancjum jest owocem pozornym róży oraz owocem jabłkowatym (np. jabłoni i gruszy).
3) Kauliforia (gr. kaulos - łodyga, flos, floris - kwiat) - jest to wyrastanie kwiatów z pąków śpiących (spoczynkowych) z pni i gałęzi. Termin ten bywa także wąsko ujmowany jako wyrastanie kwiatów tylko z pni, podczas gdy wyrastanie ich z gałęzi określane jest odrębnie jako ramifloria. Zjawisko to jest nierzadkie zwłaszcza w strefie tropikalnej, gdzie jest wynikiem adaptacji do zapylania kwiatów przez zwierzęta, które mają utrudniony dostęp do nich jeśli rozwijają się ukryte w gęstwinie liści w koronie drzewa. Dotyczy to zwłaszcza kwiatów zapylanych przez ptaki (ornitogamia), nietoperze (chiropterogamia), ale też odwiedzanych przez drobne zwierzęta wędrujące po pniach i konarach (np. gryzonie). Następujące w następstwie rozwijanie się owoców na pniach jest nazywane kaulikarpią i także wiąże się z ułatwieniem dostępu do nich niektórych zwierząt (np. małp). Kaulikarpia jest rzadkos potykana wśród drzew rosnących w umiarkowanym klimacie -w ystępuje np. u warzynka wilczełyko, judaszowca. Do przykładów licznych rodzajów i gatunków tropikalnych, u których kaulifloria ma miejsce należą: kakaowiec, chlebowiec, dzbaniwo, kalebasowe.

Źródło: Wikipedia. Judaszowiec kanadyjski - przykład rośliny u której zachodzi kaulifloria.
|
4) Kielich (łac, ang. calyx) - jest to najbardziej zewnętrzna część kwiatu, składająca się z okółka zielonych działek kielicha (łac. sepala, ang. sepals), które rozwinęły się głównie z normalnych liści podkwiatkowych. Zasadniczą funkcją kielicha jest osłanianie wewnętrznych części kwiatu. Zwykle jest zielony. U niektórych roślin kielich pełni dodatkowo funkcję powabni, mającej za zadanie przywabianie owadów. Zazwyczaj ma to miejsce w kwiatach o zredukowanych płatkach korony. Wówczas kielich pełni funkcję płatków korony i jest jaskrawo ubarwiony. Pod kielichem u niektórych roślin występuje kieliszek (epikalyx), np. u truskawki.

Źródło: Wikimedia. Pappus u różnych roślin: a - willow herb (Epilobium); b - dwie formy nasiona Thrincia hirta; c-Tmarix; d-Willow (Korzeń); e- cotton grass (Eriophorum); f-bulrush (Typha).
RODZAJE KIELICHA ZE WZGLĘDU NA JEGO TRWAŁOŚĆ TO:
- kielich odpadający. Zasadnicza funkcja kielicha kończy się z chwilą rozwoju kwiatów i wówczas u wielu roślin kielich odpada,
- kielich opadający - kielich opadający wraz z płatkami korony, dopiero po przekwitnięciu kwiatu,
- kielich trwały - pozostający po przekwitnięciu kwiatu i biorący udział w osłanianiu młodego owocu. Czasami podczas rozwoju owocu kielich znacznie rozrasta się (tzw. kielich powiększony). U niektórych roślin wiatrosiewnych przekształca się w puch kielichowy (pappus), tworzący aparat lotny umożliwiający długie unoszenie się nasion w powietrzu.

Źródło: Wikipedia. Trwały 5-działkowy kielich u pomidora.
RODZAJE KIELICHA ZE WZGLĘDU NA BUDOWĘ: Wierzba; kielich zrosłodziałkowy - jego działki są z sobą zrośnięte na całej długości, lub częściowo. W kielichu zrosłodziałkowym wyróżnia się części takie jak: rurka kielicha (najniżej położona część kielicha), zewnętrzny rąbek kielicha i gardziel kielicha; kielich wolnodziałkowy (wolny) - o wolnych, niezrośniętych z sobą działkach (np. w rzepaku).

Źródło: Wikipedia. Zrosłodziałkowy kielich lepnicy czerwonej.
Ze względu na kształt wyróżnia się wśród kielichów zrosłodziałkowych następujące typy: dwuwargowy, dzbankowaty, lejkowaty, maczugowaty, rozdęty, stożkowaty.

 |
| Źródło: Wikipedia. |
Źródło: biologydiscussion.com - formy kielicha.
--> Kieliszek (ang. łac. epicalyx) - są to występujące w kwiatach niektórych roślin dodatkowe listki znajdujące się na zewnątrz działek kielicha i zwykle do nich podobne. Występują np. u niektórych przedstawiciele rodzin różowatych (Rosaceae) i ślazowatych (Malvaceae). Obecność kieliszka jest ważną cechą diagnostyczną. We wzorze kwiatowym okółek działek kieliszka oznacza się literą E lub k (w przeciwieństwie do kielicha K).
5) Korona kwiatu (łac. corolla, ang. corolla) - jest to element kwiatu, który składa się z okółka barwnych płatków korony (petala), które stanowią powabnię dla owadów, lub innych zwierząt zapylających kwiaty. Korona stanowi wewnętrzną część okwiatu. Barwa płatków jest spowodowana występowaniem barwników w soku komórkowym (np. antocyjany). Olejki eteryczne, które nadają kwiatom zapach, występują zazwyczaj w komórkach epidermy płatków. Płatki korony składają się z rozszerzonej zwykle blaszki i wąskiego paznokcia, za pomocą którego są przytwierdzone do dna kwiatowego. Na płatkach korony u wielu roślin występują miodniki wytwarzające nektar. Czasami całe płatki przekształcone są w miodniki - tzw. listki miodnikowe. Korony mogą być: wolnopłatkowe (np. u gorczycy Sinapis); zrosłopłatkowe (np. u powoju Convolvulusa). Najczęściej występujące kształty korony zrosłopłatkowej to: dwuwargowa, dzbankowata, dzwonkowata, jednowargowa, kulista, lejkowata, lichtarzowata, talerzowata. Kształt płatków korony jest ważną cechą gatunkową prozy oznaczaniu wielu gatunków roślin. U niektórych grup roślin występują korony kwiatowe o specyficznej budowie. Np. u narcyzów występuje wewnątrz korony dodatkowy element, tzw. przykoronek, u żywokostu lekarskiego łuseczki, zwane osklepkami. U niektórych roślin (np. u orlika, ostróżki) jeden z płatków korony (czasami działka kielicha) są wydłużone tworząc tzw. ostrogę. U roślin z rodziny bobowatych płatki korony mają różną wielkość i ustawione są w specjalny sposób; górny i największy tworzy tzw. żagielek, dwa boczne tzw. skrzydełka, a dolne dwa zrośnięte z sobą tworzą łódeczkę.
 |
| Źródło: Wikipedia. Miodniki na głównym nerwie liścia śliwy. |
- Miodniki, nektarniki (łac. nectaria) - są to jedno- lub wielomomórkowe organy gruczołowe roślin wydzielające nektar. Mają one bardzo różnorodną formę, mogą być niepozorne lub okazałe i barwne. Zlokalizowane zwykle w kwiecie służą do zwabiania zwierząt, głównie owadów, dokonujących zapylania. Rośliny posiadające miodniki i wytwarzające nektar nazywane są roślinami miododajnymi. Miodniki wydzielają nektar służący do wabienia zwierząt dokonujących zapylania kwiatów. Nekrat zawiera węglowodany w różnym stężeniu, w związku z czym u jednych gatunków roślin bywa wodnisty, u innych gęsty. Różne gatunki z różną intensywnością wydzielają nektar, przy czym dla roślin korzystne jest wydzielanie ograniczonych jego ilości. Zmusza to bowiem zwierzęta zapylające do odwiedzania wielu kwiatów, co ułatwia zapylenie krzyżowe. Nektaria występują najczęściej w obrębie kwiatów (nektaria kwiatowe), czasami jednak na innych częściach rośliny; na ogonkach liściowych i głównym nerwie liści a(np. u akacji i śliwy), na liściach przykwiatowych, lub w kątach nerwów liści. Są to tzw. nektaria pozakwiatowe. W obrębie kwiatu miodniki występować mogą u podstawy słupka, na wewnętrznej stronie płatków korony czy u pdostawy płatków korony. Czasami funkcję miodników pełnią niektóre przekształcone pręciki tzw. prątniczki. Tak jest np. u dziewięciornika błotnego.
- Przykoronek (łac. corollula) - są to łuski występujące w kwiatach niektórych roślin w miejscu, gdzie blaszka płatka korony przechodzi w paznokieć. Mogą one być wolne, lub zrośnięte w rurkę, miseczkę. Tworzą charakterystyczny twór. Pełnią rolę powabni przywabiającej owady oraz ochraniają elementy rozrodcze kwiatu (pręciki i słupki). Przykoronek występuje np. u gatunków roślin w rodzinie amarylkowatych, goździkowatych i męczennicowatych.
- Osklepki (łac. fornices) - są to wyrostki (przeważnie łuskowate), zamykające wlot do gardzieli w kwiatach niektórych gatunków roślin. Występują np. w rodzinie ogórecznikowatych (Boraginaceae).
 |
| Źródło: Wikipedia. Wyraźne rurkowate przykoronki w kwiatach narcyza. |
 |
| Źródło: Wikipedia. Pięć żółtych osklepek przy wlocie rurki korony niezapominajki leśnej. |
 |
| Źródło: Wikipedia.Ostroga u ostróżki tatrzańskiej. |
- Ostroga (łac. calcar, ang. spur) - są to ostro zakończone, podługowate twory występujące w budowie morfologicznej roślin. Termin odnosi się do występującego w kwiatach niektórych roślin wyrostka u nasady płatka korony, działki kielicha lub listka miodnikowego. Jest to zamknięty na końcu wyrostek o zazwyczaj walcowatym lub stożkowatym kształcie. Jego występowanie i morfologia mają znaczenie przy oznaczaniu gatunków niektórych roślin. Ostroga występuje np. u niektórych roślin z rodziny fiołkowatych, jaskrowatych, storczykowatych. Najczęściej pełni funkcję powabni - zwabia owady. U niektórych roślin, np. u ostróżeczki polnej wewnątrz ostrogi, na jej końcu znajduje się miodnik. Mianem ostrogi określa się także ostro zakończone, wąskie wyrostki blaszki liściowej występujące u niektórych traw na granicy blaszki i pochwy liściowej. Ostrogi takie zwykle obejmują źdźbło.
 |
| Źródło: Wikipedia. Prątniczki u powojnika. |
6) Prątniczki (łac. staminodium, liczba mnoga łac. staminodia) - są to znajdujące się w kwiatach u niektórych gatunków roślin płonne pręciki, które nie mają pylników i nie wytwarzają pyłku. Spełniać mogą natomiast inne funkcje. Np. u piwonii upodabniają się one do płatków korony, pełniąc rolę powabni dla owadów zapylających kwiaty. Podobną funkcję spełniają u powojnika (Clematis), tworząc wewnątrz korony ozdobny pierścień prątniczek. Niektóre z uprawianych odmian ozdobnych powojnka kwiecistego (Clematis florida) mają wszystkie pręciki przekształcone w ozdobne prątniczki. U niektórych gatunków w rodzinie jaskrowatych prątniczki pełnią funkcję miodników. U tawułowych (rodzina różowatych) prątniczki zrastając się tworzą czerwony pierścień oddzielający pręciki od słupków. U urdzika karpackiego drobne, łuskowate prątniczki występują pomiędzy normalnymi, płodnymi pręcikami.
7) Pręcik (łac. stamen) - jest to męski organ płciowy w kwiecie wytwarzający pyłek, czyli odpowiednik męskiego gametofitu powstający z mikrospory. Pręciki są występującymi u okrytonasiennych mikrosporofilami. Składają się z nitki i główki tworzonej zazwyczaj przez dwa pylniki zaweirające po dwa woreczki pyłkowe, czyli mikrosporangia. Pyłek po dojrzeniu wysypywany jest z pękających pylników i przenoszony przez wiatr, wodę lub zwierzęta, by po osadzeniu na znamionach słupków wrosnąć i dokonać zapłodnienia. Pręciki w kwiatach poszczególnych gatunków występują w różnej liczbie, tworząc pręcikowie. Jeśli jedynymi w kwiecie organami generatywnymi są pręciki, to kwiat taki określany jest mianem pręcikowego (męskiego), gdy kwiat pozbawiony jest pręcików - określany jest mianem słupkowego (żeńskiego). U roślin nagonasiennych występują mikrosporangia zwane pylnikami na łuskowatych mikrosporofilach zwanych czasem pręcikami. Tworzą one zwykle szyszkowate twory zwane mikrostrobilami lub kwitostanami męskimi.
 |
| Źródło: Wikipedia. Główka pręcika rzodkiewnika (pęknięte pylniki z wysypującym się pyłkiem). |
MORFOLOGIA: Wyróżniane są dwa główne typy morfologiczne pręcików - pręciki blaszkowate oraz pręciki nitkowate, z możliwością wyróżniania typów pośrednich. Pręciki laminarne mają budowę zbliżoną do liścia z woreczkami pyłkowymi zwykle po stronie doosiowej, u Degenaria, Annonaceae i Humantandraceae po stronie odosiowej.

Pręciki mogą występować w różnej liczbie w kwiecie, mogą być wolne, zrośnięte ze sobą lub przyrośnięte do okwiatu lub słupkowia. Liczba pręcików, ich układ, sposób zrośnięcia jest jedną z istotnych cech diagnostycznych, analizowanych przy ustalaniu przynależności systematycznej roślin. Wszystkie pręciki w kwiecie określane są zbiorczą nazwą jako pręcikowie (łac. androeceum). Powstaje ono w kwiecie jako trzeci okółek po kielichu i koronie, a przed powstającym centralnie słupkowiem. Jeżli pręciki tworzą jeden okółek, określane są mianem haplostemon, najczęściej jednak tworzą dwa okółki. Jeśli pręciki drugiego okółka są międzyległe względem pirwszego - układ taki określany jest terminem diplostemon, jeśli pręciki drugiego okółka są nadległe pierwszemu - układ opisywany jest jako obdiplostemon. W zależności od ułożenia pręcików względem listków okwiatu wyróżniane są pręciki nadległe działkom kielicha (episepala) lub płatkom korony (epipetala). Wyjątkowo, u niektórych przedstawicieli tryurydowatych (np. Lacandonia schismatica), pręciki zajmują położenie centralne, a słupki otaczają je od zewnątrz.
 |
| Źródło: Wikipedia. Pręciki (brązowe) nadległe płatkom korony (czerwone) i pręciki (brązowe) nadległe działkom kielicha (zielone). |
 |
| Źródło: Wikipedia. |
Najczęściej pręcik u roślin okrytonasiennych składa się z nitki pręcikowej (filamentum) i główki (anthera), która jest zróżnicowana zazwyczaj na dwa pylniki (thecae) połączone płonnym łącznikiem (connectivum). Modyfikacje budowy i funkcji sprawiają, że pręciki mogą u poszczególnych gatunków być bardzo różnie wykształcone. Jednym z częściej spotykanych modyfikacji budowy pylników jest zlanie się dwóch woreczków pyłkowych w jeden. Różnice wielkości pręcików są znaczne. Na przykład u strelicji płodne pręciki osiągają 5 cm długości, a u Lepianthes (pieprzowate) tylko 0,2 mm. W zależności od położenia główki pręcika określa się ją jako wewnątrzpylną (główka zwrócona do osi kwiatowej) lub zewnątrzpylną (główka odwrócona na zewnątrz). W zależności od osadzenia główki na nitce wyróżnia się układ, w którym nitka od nasady główki wnika między pylniki w postaci łącznika (basifixed) lub łączy się z łącznikiem od tyłu główki (dorsifixed).
Modyfikacje budowy i układu pręcików w kwiecie pozwalają często na uniknięcie lub minimalizację ryzyka samozapylenia, rzadziej sprzyjają temu zjawisku. Wiele adaptacji służy zwiększeniu szans na zapylenie krzyżowe. W efekcie pręciki często różnią się długością od słupka, a także pomiędzy sobą. W kwiatach rodziny jasnotowatych (Lamiacae) znajdują się cztery pręciki, z których dwa są długie, a dwa krótkie. Takie pręciki określane są mianem dwusilnych. W rodzinie kapustowatych (Brassicaceae) cztery pręciki są długie, a dwa krótkie - określane są mianem czterosilnych. Pręciki w kwiatach przystosowanych do zapylania przez zwierzęta są często ukryte i wraz z innymi elementami kwiatu tworzą wyspecjalizowane mechanizmy, czasem dla określonego gatunku zapylacza. Przystosowaniem do wiatropylności są pręciki o długich, wystających z kwiatu i wiotkich nitkach, wytwarzające w pylnikach wielkie ilości lekkiego pyłku.
Pręciki mogąb yć ze sobą zrośnięte nitkami (np. u ślazu) lub pylnikami (np. u ostu). Zrośnięte mogą być w jedną lub kilka wiązek. U wielu bobowatych 9 pręcików jest zrośniętych, a jeden górny jest wolny. Pręciki mogą być przyrośnięte do płatków korony, czasem na całej długości nitki, tak, że pylniki są siedzące (np. u werbeny). W rodzinach: storczykowatych, kokornakowatych, toinowatych i Stylidiaceae pręciki zrośnięte ze słupkiem tworzą prętosłup.
W kwiatach klejstogamicznych występuje często redukcja pręcikowia. Przykładowo w takich kwiatach u fiołków liczba pręcików spada z 5 do 1 lub 2. Czasem zmniejsza się w takich kwiatach także liczba woreczków pyłkowych i samego pyłku. W skrajnych przypadkach zmodyfikowane pręciki przestają pełnić swoją podstawową funkcję - stają się płonne i nie wytwarzają pyłku. Tak zmodyfikowane organy są zwane prątniczkami.
ROZWÓJ I BUDOWA ANATOMICZNA: W budowie wewnętrznej główki pręcika występują zwykle cztery woreczki pyłkowe (mikrosporangia) połączone parami w pylniki zrośnięte za pomocą łącznika stanowiącego przedłużenie nitki. W kwiatach niektórych rodzin występuje jednak inna liczba woreczków pyłkowych. Tylko dwa woreczki pyłkowe występują w główce pręcika u roślin z rodzin: piżmaczkowatych, ślazowatych i rześciowatych. U niektórych gatunków z dyniowatych występują zarówno pręciki zz dwoma, jak i z czterema woreczkami pyłkowymi. W sumie gatunki, w których przypadku zarejestrowano redukcję liczby woreczków pyłkowych z czterech do dwóch, reprezentują aż 60 rodzin. Więcej niż cztery woreczki pyłkowe występują w pręcikach storczykowatych. W rodzinie goryczkowatych i gązewnikowatych w woreczkach pyłkowych powstają przegrody poprzeczne. Rozwój kwiatów oraz poszczególnych ich elementów został dobrze poznany na przykładzie Arabidopsis thaliana. W merystemie kwiatowym, podobnie jak w merystemie wierzchołkowym pędu, można wyróżnić kilka warstw (histogenów). W jednym z ujęć wyróżniane są trzy warstwy: epidermalna (L1), subepidermalna (L2) oraz rdzeń (L3). W innym, warstwy L1 i L2 określane są jako tunika, a warstwa L3 jako korpus. Związki pręcików powstają w wyniku podziałów peryklinalnych (podział peryklinalny to podział komórki, w którym nowo powstająca ściana podziałowa jest równoległa do powierzchni stycznej organu) komórek zwykle warstwy L2. U dwuliściennych elementy kwiatu zorganizowane są w czterech okółkach, których zawiązki pojawiają się kolejno podczas rozwoju merystemu kwiatowego. Okółek złożony z pręcików u A. thaliana, jest trzecim i skłąda się z sześciu pręcików, czterech środkowych (długich) i dwóch bocznych (krótkich). Morfogeneza kwiatu została podzielona na 12 etapów. Pręciki rozpoczynają swój rozwój na etapie 5. Na etapie 7. zawiązki pręcików skłądają się już z warstwy epidermalnej (L1), pojedynczej warstwy subepidermalnej (L2) oraz komórek warstwy rdzeniowej (L3), z której powstaje wiązka przewodząca. U innych gatunków cały zawiązek pręcika może powstawać niemal wyłącznie z warstwy L2. Przez pewien czas zawiązek rozwija się dzięki podziałom w obrębie merystemu szczytowego, dlaszy wzrost ma charakter interkalarny. U niektórych gatunków pojawia się także merystem brzeżny, prowadzący do powstania blaszkowej formy pręcika lub też spłaszczonej nitki. Wzrost nitki pręcikowej na długość u traw jest najszybszym takim procesem w świecie roślin - osiąga on tempo 1mm na minutę, podczas gdy wzrost organów większości roślin nie przekracza kilku mikrometrów na minutę (pędy bambusa przyrastają 0,2 mm na minutę). Górna część zawiązka pręcika ulega pogrubieniu, tworząc zaczątek pylnika. Zaczątek ten wydłuża się, a na przekroju poprzecznym przybiera kształt trapezu. W narożach pojawiają się komórki subepidermalne o dużych jądrach, które po podziale peryklinalnym tworzą dwie warstwy. Warstwa wewnętrzna to pierwotne komórki sporogenne (zwane także komórkami pramiacierzystymi pyłku), a warstwa zewnętrzna to komórki ścienne (parietalne). Antyklinalny rozrost obu warstw prowadzi do powstania czterech nabrzmień, z których powstają woreczki pyłkowe. Część pylnika położona w głębokim wcięciu tworzy łącznik. Komórki sporogenne rozwijają się w mikrosporocyty, nazywane także komórkami macierzystymi pyłku lub mejocytami męskimi. U niektórych gatunków w komórkach pramacierzystych pyłku od razu zachodzi mejoza i stanowią one bezpośrednio komórki macierzyste pyłku. U innych zachodzi w nich najpierw szereg podziałów mitotcznych, w których wyniku komórki potomne ustawiają się szeregowo, i dopiero wówczas następuje mejoza i powstają komórki macierzyste pyłku. Koncentryczne pierścienie komórek wokół komórek sporogennych stanowią mikrosporangium. Wszystkie te typy komórek istnieją już na etapie 9. rozwoju kwiatu. Mikrosporocyty uwalniane są do wnętrza woreczków pyłkowych, gdzie rozwijają się w gametofity męskie. Na etapie 12. w woreczkach pyłkowych znajdują się trzykomórkowe ziarna pyłku.
BUDOWA PRĘCIKA: 1) NITKA PRĘCIKA; 2) GŁÓWKA ZŁOŻONA Z DWÓCH PYLNIKÓW; 3) ŁĄCZNIK; 4) KOMORA PYŁKOWA.
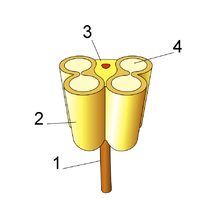
BUDOWA PRĘCIKA: 1) NITKA PRĘCIKA; 2) GŁÓWKA ZŁOŻONA Z DWÓCH PYLNIKÓW; 3) ŁĄCZNIK; 4) KOMORA PYŁKOWA.
Komórki ścienne woreczków pyłkowych po podziałach antyklinalnych i peryklinalnych tworzą trzy warstwy. Najbardziej zewnętrzna, położona bezpośrednio pod epidermą warstwa, nazywana endotecjum, składa się z komórek, w których wtórna ściana komórkowa drewnieje według określonego wzoru. Zgrubienia wtórnej ściany komórkowej tworzą usztywnione listwy, od zewnątrz cienkie, od wnętrza pylnika - zgrubiałe. W wyniku utraty w tych komórkach wody i w efekcie zjawiska kohezji, między komórkami powstają szczeliny i w końcu ściana pylnika pęka. Podłużne pęknięcie pylnika połączone z rozerwaniem endotecjum między dwoma woreczkami pyłkowymi w jego obrębie występuje u ok. 90% roślin okrytonasiennych. U wielu przedstawicieli magnoliowych oraz u oczarowatych pękn^ęcie jest na jednym z końców rozwidlone. Dwie szczeliny powstają w pylnikach rutowatych, meliowatych i niektórych jednoliściennych. U niektórych wawrzynowatych oba woreczki pyłkowe odsłaniane są osobnymi fałdami pękającego pylnika. U pewnych gatunków endotecjum ze zgrubieniami nie ma wcale. W taki przypadkach otwarcie następuje wskutek ztłuszczania, rozpuszcxzania i pękania ścian pylników. Powstają w efekcie nie szczeliny, lecz pory (np. u wrzosowatych) lub klapki (np. u smaczliwki). Przynajmniej u niektórych roślin pęknięcia i inne otwarcia pylników mogą otwierać się i zamykać w zależności od warunków pogodowych - otwierają się, gdy jest sucho, a zamykają, gdy pada deszcz. U roślin wodnych często brak endotecjum (np. u rogatka i w zanurzonych kwiatach rzęśli - w kwiatach wynurzonych tego rodzaju endotecjum powstaje).
Druga warstwa komórek zlokalizowana wewnątrz endotecjum to jednorzędowa lub wilorzędowa warstwa miękiszu. U starszych linii rozwojowych okrytonasiennych zwykle jest wielowarstwowa (np. magnoliowatych i jaskrowatych), u młodszych - cienka lub całkiem zredukowana. Komórki w niej są bogate w skrobię i tłuszcze zużywane stopniowo w miarę rozwoju pyłku. W miarę dojrzewania pylników i pyłku komórki tej warstwy są zwykle zzgniatane. Najbardziej wewnętrzna warstwa pylników, nazywana tapetum, wyściela woreczki pyłkowe i składa się z komórek wydzielniczych o dużych jądrach i gęstej cytoplazmie z licznymi rybosomami oraz rozbudowanym systemem siateczki śródplazmatycznej. Komórki te wydzielają do wnętrza pylników płyn (Zwany czasem "sokiem pylnikowym") zawierający aminokwasy, polisacharydy, enzymy i substancje wzrostowe. W wydzielinie tej rozwijają się ziarna pyłku. U licznych taksonów roślinnych (np. w rodzinie astrowatych i komelinowatych) ściany komórek tapetum ulegają rozpuszczeniu i ich protoplasty zlewają się, tworząc wielojądrowe plazmodium, w którym znajdują się ziarna pyłku. W miarę zużywania treści komórek tapetum przez rozwijający się pyłek - warstwa ta stopniowo zanika.
Ostatnim etapem rozwoju pręcika jest odwodnienie pylników i pyłku. W efekcie ziarna pyłku wchodzą w stan uśpienia i stają się sypkie, co ułatwia ich rozprzestrzenianie. W niektó®ych rodzinach ziarna ypłku pozostają złączone po cztery w tetradach (Np. u wrzosowatych i sitowatych). W przypadku wielu roślin owadopylnych pyłek opleciony jest lepką wiscyną (kitem pszczelim), ułatwiającym jego przenoszenie przez owady. Taki pyłek ma m.in. różanecznik, wierzbówka i fuksja. Pyłek z całego pylnika zleiony w tzw. pyłkowinę występuje u storczykowatych i trójeściowatych.
Proces uwalniania pyłku jest zróżnicowany. Niektóre rośliny (np. trawy) wysypują cały pyłek naraz po pęknięciu pylników, inne czynią to stopniowo. W przypadku kwiatów z wieloma pręcikami zwykle pyłek uwalniany jest sukcesywnie. Czas pylenia może różnić się w zależności od mechanizmu uwalniania pyłku oraz liczby pręcików i wynosi od kilkudziesięciu minut do kilku dni. W dominujących kwiatach przedprątnych pyłek wysypuje się krótko po rozchyleniu się listków okwiatu. W kwiatach przedsłupnych najpierw dojrzewają słupki. Po kwitnieniu pręciki są odrzucane wraz z całym kwiatem (w przypadku kwiatów niezapłodnionych) lub zwykle odpadają wraz z okwiatem podczas zawiązywania owoców.
KONTROLA GENETYCZNA ROZWOJU: Molekularne mechanizmy rozwoju kwiatów zostały poznanie głównie dzięki badaniom na Antirrhinum majus oraz Arabidopsis thaliana. Podobieństwa pomiędzy oboma gatunkami pozwoliły stwierdzić, że za genetyczną kontrolę rozwoju kwiatów odpowiada grupa genów homeotycznych [ang. homeobox - grupa genów kontrolujących rozwój morfologiczny poszczególnych części ciała początkowych stadiach rozwoju zarodkowego] należące do trzech różnych klas A, B i C. Za wykształćenie pręcików odpowiadają wspólnie geny z klasy B i C. U Arabidopsis do klasy B należą geny: APETALA3 (AP3) oraz PISTLLATA (PI), a do klasy C gen AGAMOUS (AG). Geny klasy B u Antirrhinum to DEFICIENS (DEF) oraz GLOBOSA (GLO), zaś klasy C - PLENA. Mutacje w dowolnym genie z klasy B prowadzą do wykształcenia, zamiast pręcików, owocolistków, a w przypadku mutacji w genach klasy C zamiast pręcików powstają płatki korony. Przy utracie funkcji przez geny obu klas w miejsce pręcików wykształcają się działki kielicha.
 |
| Źródło: Wikipedia. Dodatkowe płatki w odmianach pełnokwiatowych powstają ze zmodyfikowanych pręcików wyniku mutacji genów klasy c (Papaver atlanticum). |
REGULACJA HORMONALNA I ROLA GIBERELIN W ROZWOJU PRĘCIKÓW: Rozwój pręcików jest koordynowany przez fitohormony. Przy niedoborze giberelin, spowodowanym mutacją, pręciki nie wydłużają się i w efekcie powstają kwiaty męskosterylne. Ten sam efekt wywołuje niedobór jasmonianów. Gibereliny wyzwalają podczas rozwoju pręcika degradację białek DELLA. Przy niedoborze giberelin dochodzi do akumulacji tych białek, co jest bezpośrednią przyczyną powstania męskosterylnych kwiatów. Dane doświadczalne wskazują, że gibereliny również regulują syntezę jasmonianów. Zależność pomiędzy tymi hormonami jest więc hierarchiczna. Kwas jasmonowy reguluje końcową fazę wydłużania nitki dojrzewania pyłku i rozejście się brzegów pylników. Późne etapy rozwoju pręcika są jednocześnie regulowane przez auksyny, które uczestniczą także w koordynacji wcześniejszych etapów rozwoju pręcików. U mutantów z defektem syntezy auksyn pręciki powstają, anie ulegają jednak wydłużaniu i nie dochodzi do rozwoju pylników. Zwiększone ilości kwasu abscysynowego (ABA) prowadzą do wytworzenia kwiatów męskosterylnych, z nieprawidłowo rozwiniętymi pręcikami. Właściwy rozwój pręcików u mutantów z nadmiarem ABA może być przywrócony przez poddanie roślin działaniu niskich temperatur. Jako model do badań nad rolą etylenu w regulacji hormonalnej powstawania kwiatów jednopłciowych wykorzystywany jest ogórek (Cucumis sativus) . Etylen jest kluczowym hormonem na etapie powtawania zawiązków pręcików. Przy nadmiernej ilości etylenu pręciki nie wykształcają się, a kwiaty są wyłącznie żeńskie, zaś u mutanta melona z niedoborem etylenu powstają kwiaty wyłącznie męskie. Przeniesienie zmutowanego genu receptora etylenu z melona do tytoniu (Nicotiana tabacum) prowadzi do nieprawidłowego rozwoju pręcików i zmniejszenia ilości wytwarzanego pyłku.
MODYFIKACJE TRANSGENICZNE: Ponieważ do rozwoju płodnego pyłku niezbędne jest powstanie normalnej warstwy tapetum, zaburza się jego funkcje w celu uzyskania roślin męskosterylnych. Uzyskano takie transgeniczne odmiany rzepaku i tytoniu poprzez wprowadzenie do nich genów dwojakiego pochodzenia. Promotor pochodził z genu transkrybującego powstawanie białek w tapetum. Zamiast jego drugiej części wstawiano gen determinujący syntezę rybonukleaz pochodzący z bakterii (Bacillus liquefaciens) lub grzybów (Aspergillus oryzae). W efekcie uzyskano rośliny pod każdym względem normalne, ale ze zdegenerowanym tapetum. Uzyskanie roślin męskosterylnych jest jedną ze strategii mających zapobiec rozprzestrzenianiu się genów z roślin transgenicznych w populacjach dziko rosnących.
MODYFIKACJE TRANSGENICZNE: Podstawową funkcją pręcików jest wytwarzanie pyłku. W pylnikach podczas sporogenezy powstają mikrospory z komórek sporogennych. Inne komórki tworzące pylnik zapewniają dojrzewanie i ochronę pyłku oraz umożliwiają jego uwolnienie. Przez nitkę dostarczane są do pylników związki odżywcze oraz woda. Nitka zapewnia także odpowiednie ułożenie pylników w przestrzeni, umożliwiające rozprzestrzenienie się dojrzałego pyłku. U wielu roślin pręciki uzupełniają lub zastępują okwiat w roli powabni. Okazałe i barwne pręciki występują m.in. u mimoz, akacji, wielu przedstawicieli mirtowatych, ślazowatych i wawrzynkowatych. U niektórych psiankowatych pręciki uwalniają olejki aromatyczne wabiące zapylaczy. U mimozowych płonne przedłużenie nitki i łącznika tworzy twór, w którym gromadzone są substancje odżwycze, co także przyciągać ma zapylaczy. U 32,5% rodzin i 54,4% rodzajów roślin okrytonasiennych występują pręciki pozbawione pylników - prątniczki (łac. staminodium). Duże rozpowszechnienie wśród rodzajów wynika głównie z bogactwa storczykowatych z kwiatami zawierającymi prątniczki, wśród innych rodzin pręciki niewytwarzające pyłku występują u 5,9% rodzajów. Po ewolucyjnej utracie pierwotnej funkcji prątniczki zaczęły pełnić wiele nowych. Mogą one zapobiegać samozapyleniu, tworzyć sygnał wizualny lub zapachowy dla zapylaczy, ewentualnie zapewniać im nagrodę po przekształceniu w miodniki (łac. nectaria). W kwiatach Xyris prątniczki zakończone kępkami włosów umożliwiają wtórne przeniesienie pyłku na zapylacza. U Commelina stymulują pszczoły do wykonywania dodatkowych ruchów. Prątniczki mogą także otaczać i chronić przed uszkodzeniem zalążnię.
RUCHY PRĘCIKÓW: Ruchy pręcików stwierdzono w kwiatach należących do przedstawicieli różnych rodzin. Ruchy te mają na celu zwiększenie szans na sukces rozrodczy. Przemieszczanie się męskich narządów rozrodczych mające na celu odpowiednią interakcję z zapylaczami zostały stwierdzone u ożwiowatych, dziewięciornikowatych, rutowatych i nasturcjowatych. Kluczowym celem ruchów jest wpływanie na częstość kontaktu i umiejscowienie pyłku na zapylaczu. Pręciki poruszają się według określonego wzoru, niekiedy jeden po drugim lub też całymi grupami bądź też grupa po grupie. Ruchy pręcików mają charakter tigmonastii (sejsmonastii - reakcji na bodziec mechaniczny wywołanej zmianą turgoru), a bodźcem wywołującym reakcję rośliny jest dotyk zapylacza. Miejsce wrażliwe na dotyk jest ściśle określone ruch może trwać od 1,5 do 2 sekund, a jego tor jest zawsze taki sam. Ruchy wykonują także pręciki w kwiatach roślin rozmnażających się autogamicznie - w wyniku samozapylenia. Ruchy takie stwierdzono m.in. u przedstawicieli skalnicowatych, goździkowatych i kapustowatych. Mają one na celu zetknięcie pylników ze znamieniem. U niektórych roślin (np. u dziewaczka i goryczki) podczas przekwitania pylniki zostają przyciśnięte do znamienia, w taki sposób, że wysypują nań pyłek. Z kolei w kwiatach kawowca pylniki pękają jeszcze w stadium pąka kwiatowego, obsypując znamię. Rzadko u roślin występują eksplozywne mechanizmy uwalniania pyłku (np. u pokrzywy i przesiąkry).
EWOLUCJA: Wielu autorów tradycyjnie za formę bardziej prymitywną uznaje pręciki blaszkowate, kształtem zbliżone do liści, czy też płatków kwiatu. Pręciki o takiej budowie występują u roślin bazalnych grup roślin okrytonasiennych - degeneriowatych, himantandrowatych, Austrobaileyaceae i grzybieniowatych. Interpretacja, w której pręciki traktowane są jako narząd homologiczny do liści, jest rozpowszechniona w literaturze, jednakże niektórzy autorzy zwracają uwagę na obecność pręcików blaszkowatych jedynie w dużych kwiatach. W takim przypadku spłaszczenie może być jedynie adaptacją zwiększającą wytrzymałość mechaniczną. Zwraca się uwagę na brak podobieństw w budowie powstałych w wyniku modyfikacji liści owocolistków i pręcików (zarówno pojedynczych, jak i tworzących złożone struktury w wyniku ich zrastania się). Odkryto także, że rozwój pręcików przebiega odmiennie niż w przypadku liści i owocolistków, a nawet w przypadku jednej z rodzin o spłaszczonych ("prymitywnych") pręcikach - Austrobaileyaceae odkryto, że blaszkowate brzegi nitki pręcika powstają wtórnie w trakcie ich rozwoju. W niektórych pracach wskazuje się na odrębną (podobną do osiowej) genezę pręcików, podczas gdy pozostałe okółki powstać miały z liści. Równocześnie uznaje się także, że w toku ewolucji płatki korony powstały z zewnętrznych pręcików.
Genetyczny model kontroli rozwoju kwiatów został poznany w wyniku badań dwóch gatunków roślin dwuliściennych. Badania na kukurydzy potwierdziłī, że model jest wspólny dla wszystkich okrytozalążkowych. Jedynie u pierwotnych okrytonasiennych takich jak Eupomatiaceae mechanizmy regulacyjne są odmienne. Nie występują u nich działki kielicha i płatki korony, a pręciki, z któ®ych część pełni funkcję powabni, a część wytwarza pyłek, są ułożone spiralnie. Geny ABC musiały powstać jeszcze przed wykształceniem roślin okrytonasiennych. Geny z tej samej rodziny zostały wykryte u mszaków, paprotników, a nawet u siostrzanej wobec roślin lądowych grupy glonów ramienicowych (Charophyceae). Ewolucja tych genów, w tym wielokrotne duplikacje, doprowadziła do powstania współcześnie występujących elementów kwiatów. Na etapie wyodrębniania magnolilidów, w wyniku duplikacji doszło do powstania dwóch linii paleoAP3 i PI. Gen paleoAP3 w wyniku duplikacji dał początek liniom euAP3 oraz TM6. Obie te linie obecne są u dwuliściennych właściwych.
 |
| Źródło: Wikipedia. Pręciki w kwiecie Nymphaea odorata - - zewnętrzne spłaszczone, wewnętrzne na cienkich nitkach. |
Do trendów ewolucyjnych dotyczących pręcików należy zmiana ich układu ze spiralnego w okółkowy i zmniejszanie się ich liczby. Za prymitywniejsze uważa się pręciki o pylnikach osadzone nasadą na nitce z masywnym łącznikiem, za młodsze - pylniki z cienkim łącznikiem łączącym się od tyłu z nitką. W młodszych grupach okrytonasiennych proporcjonalnie większy jest też udział tkanki archesporialnej w budowie pylnika. Uznaje się to za trend prowadzący do specjalizacji tego organu do funkcji rozrodczej, podczas gdy u bazalnych grup okrytonasiennych pręciki pełniły także w kwiecie funkcję ochronną i powabni. W wielu młodszych grupach wyspecjalizowane pręciki ponownie w wielu przypadkach zaczęły wykazywać różne adaptacja zwiększające sukces rozrodczy - wabiąc zapylaczy atrakcyjnym wyglądem, zapachem lub nagrodą w postaci pożywienia. Ewolucyjne uzyskiwanie dodatkowych funkcji związane jest przede wszystkim z utratą funkcji podstawowej i powstaniem prątniczków. Elastyczność ich funkcji pozwala na szybką ewolucję. U licznych gatunków prątniczkom nie można przypisać żadnej funkcji. Chociaż takie struktury powinny szybko zanikać w wyniku ewolucji, szczególnie że mogą one utrudniać zapylenie, występują często, co prawdopodobnie jest etapem przejściowym w uzyskiwaniu nowej specjalizacji.
HISTORIA BADAŃ: Wiedzę o skutkach zapylania kwiatów znano od dawnych czasów, odkąd zaczęto uprawiać figowca i palmę daktylową. Sztuczne zapylanie palmy daktylowej traktowano jako obrzęd tajemniczy i religijny. Teofrast z Eresos (-370-285 BC) opisał istnienie osobników palmy daktylowej wydających owoce, i innych, które tym pierwszym w tym "pomagają". Albert Wielki opisał pręciki jako grudki z żółtym pyłem, które "znajdują się w kwiatach prawie wszystkich roślin, raz w mniejszej, raz w większej ilości". Pręciki opisane zostały wraz z podziałem na główkę i nitkę przez profesora Joachima Junga z Hamburga (1587-1657). Pręciki jako męskie organy płciowe u roślin zidentyfikowane zostały po raz pierwszy przez Rudolfa Jakoba Camerariusa (1665-1721) w dziele "De sexu plantarum epistola" w 1694 roku. Oparł się on na wnioskach z obserwacji różnych gatunków roślin dwu- i jednopiennych oraz obupłciowych, jednak sposoby zapylania i zapładniania pozostawił do wyjaśnienia tym "którzy mieć będą wzrok bystrzejszy od wzroku rysia". Rola pręcików dowiedziona została w XVIII wieku przez takich badaczy jak: Johann Gottlieb Gleditsch (1714-1786), Joseph Gottlieb Kolreuter (1733-1806) oraz Karol Linneusz (1707-1778), który na podstawie budowy pręcikowia w kwiatach zbudował swój seksualny system klasyfikacyjny roślin. Pierwszych 10 klas obejmowało rośliny o rosnącej liczbie pręcików od 1 do 10, o równej długości. Kolejne 10 klas obejmowało roślny o pręcikach o nierównej długości, zrośniętych w różnym stopniu z sobą lub innymi organami kwiatu. Ostatnie cztery klasy rośliny objęły rośliny jednopienne, dwupienne, mieszane dwupienne i obupłciowe oraz zarodnikowe.
W 1793 roku ukazała się "teoria kwiatów" Christiana Konrada Sprengla (1750-1816) tłumacząca na bazie szczegółowych obserwacji kilkuset gatunków biologię kwiatów, w tym rolę pręcików. Jego teza "wydaje się, że rośliny nie chcą być zapylane własnym pyłkiem" stała się przedmiotem badań Karola Darwina (1809-1882), któreg oprace wraz z rozwojem teorii ewolucji, spowodowały dociekanie na temat różnorodnych "przystosowań" kwiatów. Dzięki zastosowaniu mikroskopu w XIX wieku szczegółowo poznano przebieg prawidłowej morfogenezy kwiatu. OBszerną pracę na ten temat opublikował w roku 1857 Jean-Baptiste Payer (1818-1860). Kluczową pracę dla poznania rozwoju pylników i pyłku opublikował w 1873 roku duński botanik Johannes Eugenius Warming (1841 - 1924). Na przełomie XIX i XX wieku rozwinęła się ekologia kwiatów i badania eksperymentalne, od połowy XX wieku - także nad genetycznymi podstawami biologii i ekologii kwiatów.
Odkrycia Gregora Mendla (1822-1884) zapoczątkowały poznawanie wariantów genetycznych budowy kwiatów. Opisano kwiaty, w których jeden typ elementu zastępował inny. W 1967 roku Sipra Guha i S.C. Maheshwari uzyskali z kultury pylników bielunia (rodzaju bylin lub niewielkich drzew z rodziny psiankowatych, na które składa się około 12-15 gatunków) rośliny haploidalne. Ponieważ każdy gen takiego osobnika uwidacznia się w fenotypie - rośliny uzyskiwane z kultur pyłkowych i pylnikowych zyskały ogromne znaczenie w hodowli roślin. Wraz z rozwojem technik badawczych poznawano kolejne mutanty homeotyczne, a następnie geny warunkujące rozwój komórek merystemu w elementy kwiatu w zależności od lokalizacji w merystemie. W roku 1991 w czasopiśmie Development Bowman, Smyth i Meyerowitz opisali model ABC zakładający istnienie trzech zachodzących na siebi epól o swoistej regulacji genetycznej, w którym o rozwoju pręcików decydowały geny AP3/PI i AG. Model został spopularyzowany przez pracę przeglądową Enrico Coena i Elliota Meyerowitza opublikowaną w tym samym roku w Nature. Ich zespół jednocześnie dokonał podobnych ustaleń dla Antirrhinum majus. Prostota modelu oraz wzrost wartości wiedzy o rozwoju kwiatów, szybko doprowadził do imieszczenia jego opisu w podręcznikach akademickich, a nawet do wprowadzenia go do treści nauczonych w niektórych krajach w szkołach średnich.
 |
| Źródło: Wikipedia. Tablica przedstawiająca organy płciowe typowe dla 24 klas wyróżnionych w systemie klasyfikacyjnym Linneusza z Systema Naturae. |
MODEL ABC:
 |
| Źródło: Wikipedia. |
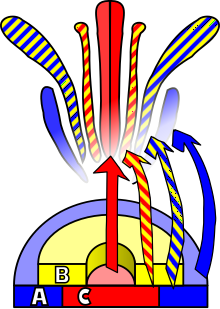
Dwie dekady temu, dzięki badaniom nad Arabidopsis thaliana i Anthirrhinum majus, E. Coen oraz E. Meyerowitz sformułowali model obrazujący genetyczną regulację powstawania określonych struktur w obrębie kwiatu, tzw. system ABC. Zakłada on zależność tożsamości kolejnych elementów budujących kwiat od interakcji trzech klas genów homeotycznych, dając w efekcie charakterystyczny dla danego okółka fenotyp. Zgodnie z zaproponowanym modelem za powsanie działek kielicha odpowiada ekspresja genów klasy A w najbardziej zewnętrznym okółku, płatków korony - współdziałanie genów klasy A i B, pręcików - jednoczesna ekspresja genów należących do klas B i C, słupków zaś - ekspresja wyłącznie genów klasy C w wewnętrznym okółku kwiatu. U rośliny modelowej Arabidopsis wyizolowano następujące geny należące do klasy A: APETALA1 (AP1) i APETALA2 (AP2), do klasy B: APETALA 3(AP3) i PISTILLATA (PI); do klasy C: AGAMOUS (AG) oraz SHATTERPROOF (SHT). Kolejne badania ukierunkowane na poznanie modelu ABC przyczyniłī się do jego rozszerzenia o kolejne klasy genów: D oraz E. To rozbudowanie modelu pozwoliło na zobrazowanie działania genów należących do klas ABC na poziomie tetramerycznych pakietów białkowych. Ekspresja genów klasy D odpowiada za prawidłowe wykształcenie zalążni, natomiast geny klasy E współdziałają z pozostałymi genami modelu, a ich produkty białkowe są niezbędnym czynnikiem nadającym pełną funkcjonalność tetramerycznym pakietom. Prawie wszystkie geny modelu ABC (z wyjątkiem AP2) należą do rodziny genów typu MADS. Przynależność ta opiera się na wspólnym planie budowy genów, czyli obecności domen: konserwatywnej MADS, łącznikowej I, charakterystycznej dla roślin domeny K oraz terminaalnej C. Rodzina MADS występuje nie tylko u roślin, ale również u grzybów oraz zwierząt. Model ABC został początkowo zaproponowany dla roślin dwuliściennych i tylko na nich był szeroko badany Okazał się być jednak bardzo dobrym narzędziem do analizy budowy kwiatu również u roślin jednoliściennych. Badania prowadzone na ryżu, pszenicy czy kukurydzy potwierdziły obecność i zasadę działanie genów należących do modelu ABC w homologicznych do roślin dwuliściennych elementach kwiatu. Ciekawych wyników dostarczyły badania nad kwiatami storczyków, gdzie nie tylko kombinacje poszczególnych klas genów odpowiadają za tożsamość kolejnych okółków, ale również obecność paralogicznych genów w obrębie klasy może zdecydowanie zmieniać fenotyp danej struktury. Na podstawie działania modelu ABC przedstawiono liczne hipotezy wyjaśniające powstanie wielu unikatowych struktur w obrębie kwiatu, szczególnie wśród pierwotnych dwuliściennych. Jedna z nich, tzw. "sliding boundary", pokazuje mechanizm przesunięcia granic ekspresji genów, którego efektem jest brak zróżnicowania okwiatu i tej grupy roślin oraz który wyjaśnia pochodzenie zjawiska petalodii obserwowanego np. u Magnolia stellata. Poznane zostały genetyczne podstawy budowy takich elementów jak staminodia (przekształcone w powabnię pręciki) czy kalyptry (struktury zamykającej elementy kwiatu w pąku). Zidentyfikowanie genów należących do rodziny MADS u roślin nasiennych, chów, paproci, a nawet u glonów, sugeruje bardzo wczesne ewolucyjnie powstanie genów należących do modelu ABC. Wielokrotne duplikacje genomu w trakcie ewolucji roślin przyczyniały się do zmian w morfologii kwiatu. Zmianom ulegała funkcja genu oraz miejsce i czas jego ekspresji. Śledząc kolejne duplikacje genów można zauważyć ich bezpośrednie odzwierciedlenie w drzewie filogenetycznym roślin, np. pierwsza znana duplikacja genów klasy B miała miejsce w momencie wyodrębnienia się ze wspólnego drzewa magnoliidów, czyli pierwotnych dwuliściennych. Model ABC opisuje jeden z mechanizmów rozwoju rośliny współdziałając z innymi programami, przyczynia się do powstania obserwowanej różnorodności morfologicznej kwiatów.
 |
| Źródło: biologyexams4u |
Model ABC zakłada zależność tożsamości kolejnych elementów kwiatów kwiatu od interakcji trzech grup genów homeotycznych, czyli takich, których ekspresja w określony miejscu i czasie reguluje aktywność innych genów na zasadzie kaskady, dając w efekcie charakterystyczne dla danego okółka fenotyp. Zgodnie z modelem każda z klas genów ulega ekspresji tylko w dwóch sąsiadujących ze sobą okółkach, w których tożsamość elementów z kolei definiowana jest przez interakcje pomiędzy tymi klasami. Według modelu, ekspresja genów klasy A w najbardziej zewnętrznym (pierwszym) okółku powoduje wykształcenie działek kielicha. Geny klasy A oraz B działające jednocześnie w drugim okółku odpowiadają za tożsamość płatków korony. W kolejnym, trzecim okółku, geny klasy A są nieaktywne ze względu na antagonizm w stosunku do genów klasy C, które to w interakcji z genami klasy B odpowiadają za fenotyp pręcików. W najbardziej wewnętrznej części kwiatu (czwarty okółek) powstają słupki jako efekt ekspresji genów wyłącznie klasy C. Antagonizm genów klasy A i C został potwierdzony w badaniach nad mutantami, u których brak ekspresji genów klasy A w pierwszym okółku powoduje wykształcenie słupka zamiast działek, ze względu na ekspresję nieblokowanych genów klasy c.
U Arabidopsis genami należącymi do klasy A są: APETALA1 (AP1) i APETALA2 (AP2). Efektem fenotypowym mutacji w obrębie tej klasy genów jest brak wykształconego okwiatu. Wskutek przesunięcia ekspresji genów klasy C na obszar nieaktywnych genów klasy A, na miejscu działek kielicha wytwarzane są słupki, natomiast zamiast płatków korony pojawia się okółek pręcików. Pozostałe wewnętrzne okółki kwiatu pozostają niezmienione. W trakcie ontogenezy kwiatu gen AP2 pełni trzy kluczowe role: współdziałając z genami APETALA1, LEAFY oraz CAULIFLOWER odpowiada za rozwój merystemu kwiatowego, z genami należącymi do modelu ABC odpowiada za tożsamość elementów okwiatu oraz kontroluje czasową i przestrzenną aktywność innych genów, biorących udział w organogenezie kwiatu. Do klasy B należą APETALA3 (AP3) oraz PISTILLATA (PI) natomiast do genów klasy C zalicza się AGAMOUS (AG) oraz SHATTERPROOF (SHT). Mutacja w obrębie genów klasy B skutkuje wykształceniem kwiatów zbudowanych z dwóch okółków działek, z powodu braku interakcji genów klas A i B oraz dwóch okółków słupków, natomiast efektem fenotypowym mutacji w obrębie genów klasy C są kwiaty pozbawione elementów generatywnych, czyli zbudowane wyłącznie przemiennie rozmieszczonych okółków działek kielicha oraz płatków korony. U Antirrhinum przedstawiecielami modelu ABC są geny: SQUAMOSA (SQUA) należący do klasy A, GLOBOSA (GLO) I DEFICIENS (DEF) należące do klasy B oraz gen PLENA z klasy C. Mutacje w obrębie każdej z klas genów mają taki sam skutek, jak u Arabidopsis.
- BUDOWA GENÓW TYPU MADS: Pierwsze badania nad sekwencjami genów DEF i AG wykazały wysoką homologię do domeny wiążącej DNA genu MCM1, odpowiadającego za regulację zmiany typu płciowego u drożdży oraz genu SRF, kodującego czynnik odpowiedzi na surowicę krwi u zwierząt. Z akronimu tych czterech genów: MCM1, AG, DEF, SRF powstała nazwa domeny MADS-box oraz nowej rodziny genów, posiadającej tę domenę w swojej budowie, tzw. rodziny genów typu MADS. Do tej grupy zaliczane są wszystkie znane geny wchodzące w skład modelu ABC, z wyjątkiem AP2. Ich cechą charakterystyczną jest obecność czterech domen: konserwatywnej dla całej rodziny domeny MADS, wiążącej DNA w homo- lub heterodimery, domeny łącznikowej - I oraz dwóch domen charakterystycznych dla podgrupy typu MIKC występującej u roślin: domeny K oraz domeny terminalnej C. Struktura białkowego produktu ekspresji domeny K jest podobna w budowie do keratyny, czyli zbudowana jest z amfipatycznych alfa-helis białkowych, uformowanych dzięki specyficznie rozmieszczonym hydrofobowym aminokwasom, pełniącym istotną funkcję przy dimeryzacji. Domena terminalna C odpowiada natomiast za tworzenie struktur trzecio- i czwartorzędowych. Rodzina MADS jest charakterystyczna nie tylko dla roślin okrytonasiennych. Geny do niej zaliczane zostały wyizolowane również z roślin nagonasiennych, a także z niższych roślin lądowych: paproci, widłaków, mszaków i glonów. Gen APETALA2, podobnie jak inne geny homeotyczne, koduje białko pełniące funkcje czynnika transkrypcyjnego, jednak nie posiada w swojej strukturze domeny MADS. Jego wyróżniającym elementem jest domena kodująca region umożliwiający przyłączenie się do DNA genu docelowego, czyli tzw. motyw AP2 lub RAP2 (z ang. related to AP2 - związany z AP2). U Arabidopsis zsekwencjonowano 12 genów o podobnej budowie i połączono je we wspólną rodzinę RAP2.
Więcej informacji znajdziesz w pracy pt. "ABC Architektury kwatu - ABC OF FLOWER ARCHITECTURE" M. Turczyn - Zakład Morfologii i Rozwoju Roślin, Insytut Biologii Roślin, Uniwersytet Wrocławski. Powyższe informacje zaczerpnięte zostały z owej publikacji.
Pomimo, że kwiaty często charakteryzują się bardzo złożoną budową i rozmaitymi typami symetrii, to najprostsze kwiaty obupłciowe mają symetrię promienistą i można w nich wyróżnić cztery koncentryczne okółki (wymienione w kolejności od brzegu kwiatu do jego środka: działek kielicha, płatków, pręcików oraz owocolistków. Tożsamość określonych części kwiatu jest określona poprzez ekspresje genów homeotycznych, decydujących o położeniu poszczególnych okółków. W ramach tzw. modelu ABC zakłada się obecność trzech las genów homeotycznych (są to odpowiednio klasy A, B, C). Ekspresja każdej z nich zachodzi w dwóch sąsiadujących ze sobą okółkach. Odkryć tych dokonano, badając fenotypy mutantów homeotycznych o zaburzonym układzie okółków w kwiecie. W przypadku Arabidopsis thaliana, znanej rośliny modelowej w badaniach genetycznych wykryto co najmniej pięć genów homeotycznych odpowiedzialnych za rozwój kwiatu.
 |
| Źródło: Wikipedia Słupek tulipana. |
8) Słupek, słupkowie (łac. pistyllum, ang pistil) - jest to żeński organ płciowy w kwiecie okrytonasiennych. Jest zbudowany ze zrośniętych ze sobą lub wolnych owocolistków (carpellae), które są zmodyfikowanymi liśćmi - najprawdopodobniej utworami homologicznymi do makrosporofili). Słupki zajmują zawsze centralne miejsce w kwiecie, gdzie mogą występować pojedynczo lub w większej liczbie. Słupkowie jest zbudowane z jednego lub z kilku owocolistków (kwiat jednoowocolistkowy, kwiat monoginiczny). W tym drugim przypadku wyróżnia się słupkowie wolnoowockowe (apokarpiczne), gdy każdy owocolistek zrasta się brzegami i tworzy odrębny słupek, lub słupkowie zrosłoowockowe (synkarpiczne), gdy owocolistki zrastają się w jeden słupek. W tym ostatnim przypadku w zależności od tego, czy owocolistki zrastają się brzegami, czy też brzegi zaginają do środka słupka - zalążnia jest jedno- lub wielokomorowa. W zależności od liczby owocolistków budujących słupek, wyróżnia się słupki jedno-, dwu-, trój- lub wielokrotne, któr emogą mieć od jednej do wielu komór w zalążni. Morfologicznie słupek skłąda się z części dolnej - zalążni (ovarium), która na górze zwęża się w szyjkę słupka i zakończona jest znamieniem. Czasem (np. u tulipana) nie wyodrębnia się szyjka - mówimy wtedy o znamieniu siedzącym. W komorach zalążni słupka znajdują się zalążki wyrastające na łożyskach.
 |
| Źródło: Wikipedia. Budowa słupka. |
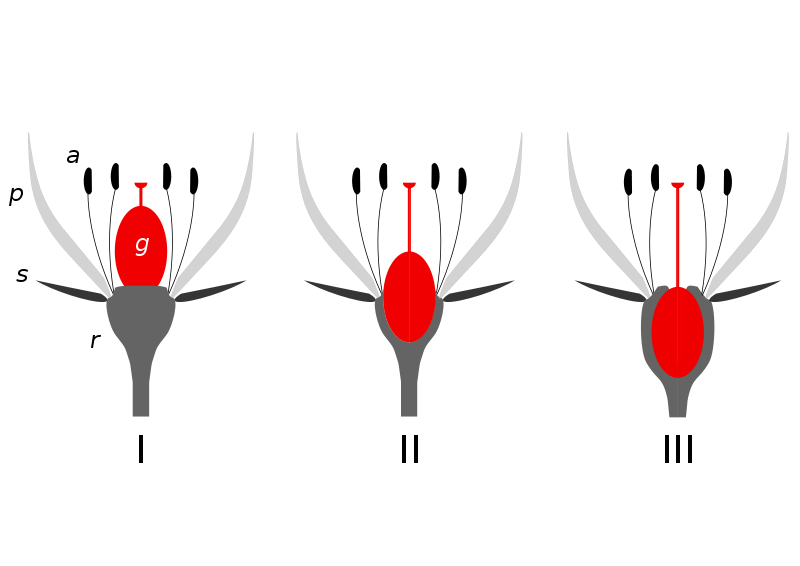
Słupki mogą być górne, pośrednie, półdolne lub dolne w zależności od położenia w stosunku do dna kwiatowego (oś kwiatowa).
SŁUPEK GÓRNY - osadzony jest luźno na wypukłym dnie kwiatowym, a pozostałe elementy kwiatu są osadzone poniżej zalążni słupka - jest to tzw. kwiat dolny (hypogyniczny), np. w kwiecie ziemniaka. Jeżeli dno kwiatowe jest wklęsłe i obrasta zalążnię, a pozostałe elementy kwiatu są wyniesione ponad nią, jest to słupek dolny, a kwiat górny (epigyniczny), np. u ogórka. W przypadku pośrednich form, np. gdy zalążnia osadzona jest swobodnie na płaskim lub wklęsłym dnie kwiatowym, nie zrastając się z nim bokami, mówimy o słupku pośrednim i kwiecie kołozalążniowym (perygynicznym). Po zapyleniu i zapłodnieniu słupek przekształca się w owoc, w którym znajdują się nasiona.
BUDOWA ANATOMICZNA:
(A) ZNAMIĘ - znamię zbudowane jest z tkanki zwanej transmisyjną (stygmatoidalną), która tworzy także połączenie z wnętrzem zalążni. Pokryte jest epidermą o charakterze gruczołowatym, czasem opatrzoną brodawkami lub włoskami. Czasem tkanka wydzielnicza może znajdować się pod epidermą. W takim przypadku wydzieliny gromadzą się najpierw w przestworach międzykomórkowych, a następnie wydostają się na zewnątrz przez luźno ułożone komórki skórki oraz pęknięcia w obrębie kutykuli.
(B) SZYJKA SŁUPKA - szyjka słupka ma wzrost interkalarny (powodowany przez merystem interkalarny), po którym następuje wydłużanie się komórek. Długość szyjki zależy od liczby podziałów poprzecznych oraz intensywności wydłużania się komórek. Podstawową tkanką budującą szyjkę jest tkanka miękiszowa. W szyjce znajduje się tkanka stygmatoidalna, podobna do tkanki znamienia. W zależności od ułożenia tej tkanki wyróżnia się szyjki otwarte i zamknięte. W typie otwartym kanał wewnątrz szyjki wyścielony jest (całkowicie lub częściowo) tkanką stygmatoidalną. W typie zamkniętym kanał jest wypełniony tkanką stygmatoidalną, której ściany komórkowe często ulegają pęcznieniu i śluzowaceniu. W przypadku słupka synkarpicznego, z kilkoma łożyskami, tkanka stygmatoidalna może tworzyć w szyjce kilka pasm, z których każde prowadzi do innego łożyska. Rozmieszczenie tkanki stygmatoidalnej w szyjce i zalążni ukierunkowuje rosnącą łagiewkę pyłkową tak, aby trafiła do zalążka (wynika to z faktu, że łagiewka nie jest w stanie przerastać pustych przestrzeni).
(C) ZALĄŻNIA - budowa anatomiczna ściany zalążni jest analogiczna do budowy liścia. Od zewnątrz znajduje się epiderma zewnętrzna, następnie miękisz ściany zalążni (mezofil) i epiderma wewnętrzna od strony komory zalążni. W skórce wewnętrznej mogą się znajdować aparaty szparkowe. W mezofilu każdego owocolistka w obrębie zalążni znajdują się wiązki przewodzące, najczęściej trzy, zbudowane z protoksylemu i protofloemu. W zalążni wykształca się łożysko - miejsce wykształcania się i rozwoju zalążków.
9) Warżka (łac. labellum) - jest to jeden z listków wewnętrznego okółka okwiatu roślin storczykowatych. W przeciwieństwie do dwóch podobnych do siebie listków bocznych tego okółka, warżka z reguły wyróżnia isę odmiennym kształtem i czasem barwą (rzadko bywa podobna do listków bocznych, np. u tajęży Goodyera - rodzaju roślin z rodziny storczykowatych, których typowym gatunkiem jest tajęża jednostronna). Zdarza się, że jest zredukowana i niemalże niewidoczna (np. w rodzaju disa Disa), jednakże na ogół jest to najbardziej efektowna i rzucają się w oczy część kwiatu.
Kształty warżek są różnorodne. Mogą mieć postać prostej płytki, mogą być zwinięte trąbkowato, wywinięte w kształt wargi lub wydęta na kształt trzewika. U niektórych gatunków warżka jest dwuczłonowa, u innych trójdzielna. W tych składających się z dwóch części, część wierzchołkowa, zwykle rozszerzona i często wcinana to epichill (zwykle ubarwiony podobnie do bocznych listków okwiatu), część dolna połączona z prętosłupem określana jest natomiast jako hypochil. Część dolna bywa zwinięta i u podstawy zaopatrzona w różnej długości, zakrzywioną ostrogę. U storczyków, u których wyróżnia się jeszcze część środkową warżki zwaną jako mezochil opisuje się w jej obrębie boczne skrzydełka (pleuridia) i płat środkowy (mesidium). Część środkowa wskazując drogę do wnętrza kwiatu ma często bardzo oryginalną budowę, jej brzegi bywają silnie powcinane lub falisto powyginane, opatrzona bywa w różnorodne wyrostki lub włoski. Warżki wydęte i działające jak pułapki na owady składają się z części nasadowej zwanej paznokciem (unquis). Otwór prowadzący do wnętrza warżki to ostium. O ile od zewnątrz warżka jest zwykle gładka i błyszcząca, to wewnątrz, zwłaszcza blisko nasady bywa silnie owłosiona. U niektórych gatunków warżka bywa wrażliwa na dotyk, także po dotarciu do jej wnętrza owada - zamyka się i przytrzymuje go zwiększając prawdopodobieństwo zapylenia. Budowa warżki jest ważną cechą taksonomiczną, pomocną przy oznaczaniu gatunków roślin z rodziny storczykowatych.
10) Osadnik (dno kwiatostanowe) - jest to silnie skrócona, rozszerzona, spłaszczona lub wypukła oś kwiatostanu, na której osadzone są kwiaty pojedyncze. Występuje w kwiatostanach typu główka oraz koszyczek.
SŁUPEK GÓRNY - osadzony jest luźno na wypukłym dnie kwiatowym, a pozostałe elementy kwiatu są osadzone poniżej zalążni słupka - jest to tzw. kwiat dolny (hypogyniczny), np. w kwiecie ziemniaka. Jeżeli dno kwiatowe jest wklęsłe i obrasta zalążnię, a pozostałe elementy kwiatu są wyniesione ponad nią, jest to słupek dolny, a kwiat górny (epigyniczny), np. u ogórka. W przypadku pośrednich form, np. gdy zalążnia osadzona jest swobodnie na płaskim lub wklęsłym dnie kwiatowym, nie zrastając się z nim bokami, mówimy o słupku pośrednim i kwiecie kołozalążniowym (perygynicznym). Po zapyleniu i zapłodnieniu słupek przekształca się w owoc, w którym znajdują się nasiona.
BUDOWA ANATOMICZNA:
(A) ZNAMIĘ - znamię zbudowane jest z tkanki zwanej transmisyjną (stygmatoidalną), która tworzy także połączenie z wnętrzem zalążni. Pokryte jest epidermą o charakterze gruczołowatym, czasem opatrzoną brodawkami lub włoskami. Czasem tkanka wydzielnicza może znajdować się pod epidermą. W takim przypadku wydzieliny gromadzą się najpierw w przestworach międzykomórkowych, a następnie wydostają się na zewnątrz przez luźno ułożone komórki skórki oraz pęknięcia w obrębie kutykuli.
(B) SZYJKA SŁUPKA - szyjka słupka ma wzrost interkalarny (powodowany przez merystem interkalarny), po którym następuje wydłużanie się komórek. Długość szyjki zależy od liczby podziałów poprzecznych oraz intensywności wydłużania się komórek. Podstawową tkanką budującą szyjkę jest tkanka miękiszowa. W szyjce znajduje się tkanka stygmatoidalna, podobna do tkanki znamienia. W zależności od ułożenia tej tkanki wyróżnia się szyjki otwarte i zamknięte. W typie otwartym kanał wewnątrz szyjki wyścielony jest (całkowicie lub częściowo) tkanką stygmatoidalną. W typie zamkniętym kanał jest wypełniony tkanką stygmatoidalną, której ściany komórkowe często ulegają pęcznieniu i śluzowaceniu. W przypadku słupka synkarpicznego, z kilkoma łożyskami, tkanka stygmatoidalna może tworzyć w szyjce kilka pasm, z których każde prowadzi do innego łożyska. Rozmieszczenie tkanki stygmatoidalnej w szyjce i zalążni ukierunkowuje rosnącą łagiewkę pyłkową tak, aby trafiła do zalążka (wynika to z faktu, że łagiewka nie jest w stanie przerastać pustych przestrzeni).
(C) ZALĄŻNIA - budowa anatomiczna ściany zalążni jest analogiczna do budowy liścia. Od zewnątrz znajduje się epiderma zewnętrzna, następnie miękisz ściany zalążni (mezofil) i epiderma wewnętrzna od strony komory zalążni. W skórce wewnętrznej mogą się znajdować aparaty szparkowe. W mezofilu każdego owocolistka w obrębie zalążni znajdują się wiązki przewodzące, najczęściej trzy, zbudowane z protoksylemu i protofloemu. W zalążni wykształca się łożysko - miejsce wykształcania się i rozwoju zalążków.
- Łożysko (placenta) - jest to miejsce wykształcenia się i rozwoju zalążków w zalążni słupka. Zalążki przymocowują się do niego za pomocą sznureczka (funiculus), przez który wnikają do zalążki wiązki przewodzące. Łożysko może powstawać w miejscu zrośnięcia isę brzegów lub powierzchni owocolistków synkarpicznych (brzeżnie - marginalnie) lub na wewnętrznej powierzchni owocolistków apokarpicznych (powierzchniowo - laminarnie). W słupkowiu synkarpicznym łożyska dzielą się na: KĄTOWE - kiedy łożyska wykształcają się w kątach komór zalążni; ŚCIENNE (PARIETALNE) - kiedy łożyska występują na brzegach niepełnych przegród, powstających na skutek zaginania się brzegów owocolistków do środka zalążni; ŚRODKOWE (CENTRALNE) - kiedy łożysko występuje w postaci centralnie położonej kolumny, bez kontaktu ze ścianą zalążni. Odmianą łożyska centralnego jest łożysko bazalne - wykształcające się u podstawy zalążni.
 |
| Źródło: Wikipedia. Przekrój przez kwiat kokoryczki - widoczna jest zalążnia z 6 zalążkami. |
*Zalążnia (ovarium) - to rozszerzona, dolna część słupka kwiatowego (pistillum) u roślin okrytonasiennych. Powstaje w wyniku zrośnięcia jednego lub większej liczby owocolistków, tworzących ściany komory zalążniowej. Zalążnia w słupku zbudowanym z jednego owocolistka jest zawsze jednokomorowa. Jeśli owocolistków jest więcej, ale zrastają się brzegami - także tworzą komorę pojedynczą. Jeśli brzegi owocolistkó∑ wchodzą do wnętrza, a ściany zalążni powstają w wyniku zrastania się ich zewnętrznych części to zalążnia może być wielokomorowa lub może zawierać niepełne przegrody. Jeśli to nie brzegi owocolistków, a wybujałe tkanki wyrastające z ich ścian dzielą komorę zalążni - nazywane są one fałszywymi przegrodami (np. u kapustowatych). Zalążnia utworzona z więcej aniżeli jednego owocolistka bywa nazywana cenokarpem. W wyniku zrastania się owocolistków (zarówno pojedynczych, jak i wielu), ich górna powierzchnia staje się wewnętrzną ścianą zalążni. Nierzadko owocolistki zrastają się w dolnej części słupka, tworzącej zalążnię, podczas gdy wyżej rozchylają się tworząc odrębne szyjki i znamiona. Na wewnętrznych ścianach owocolistków znajduje się jeden lub wiele zalążków. Powstawać mogą albo na brzegach owocolistków (submarginalnie) albo na ich powierzchni (laminalnie) z wyniosłości zwanej łożyskiem (placenta), przez które przechodzą wiązki przewodzące. U niektórych grup systematycznych roślin (u sandałowców) następuje daleko posunięta redukcja budowy i integracja zalążków z zalążnią, np. u gałecznicowatych redukcja integumentów i łożyska prowadzi do tego, że komórki macierzyste makrospor osadzone są po prostu w ścianie zalążni.
W zależności od umieszczenia względem brzegów owocolistka i liczby komór zalążni wyróżnia się następujące sposoby osadzenia zalążków: brzeżne - rozwijają się na brzegach owocolistków w zalążni jednokomorowej; kątowe - wyrastają na brzegach owocolistków w zalążni wielokomorowej; ścienne - rozwijają się na wewnętrznej ścianie zalążni; osiowe - znajdują się na kolumience wyrastając z dna zalążni jednokomorowej, powstałej ze zrośnięcia dolnych (nasadowych) brzegów owocolistków.
 |
| Źródło: Wikipedia. Położenie zalążków w zalążni: a) ścienne, b) kątowe, c) osiowe. |
W zależności od położenia zalążni względem dna kwiatowego wyróżnia się słupek górny, górny kołozalążniowy lub dolny. W czasie zapylania kwiatu łagiewka pyłkowa rośnie na wewnętrznej powierzchni zalążni, zwykle wzdłuż łożyska, dochodząc do wierzchołka zalążka. Z chwilą zapłodnienia zalążek rozwija się w nasiono, a zalążnia bierze udział w tworzeniu owocu - z jej ścian powstaje owocnia.
 |
| Źródło: Wikipedia. 1 - słupek górny, 2 - słupek górny kołozalążniowy, 3 - słupek dolny. |
9) Warżka (łac. labellum) - jest to jeden z listków wewnętrznego okółka okwiatu roślin storczykowatych. W przeciwieństwie do dwóch podobnych do siebie listków bocznych tego okółka, warżka z reguły wyróżnia isę odmiennym kształtem i czasem barwą (rzadko bywa podobna do listków bocznych, np. u tajęży Goodyera - rodzaju roślin z rodziny storczykowatych, których typowym gatunkiem jest tajęża jednostronna). Zdarza się, że jest zredukowana i niemalże niewidoczna (np. w rodzaju disa Disa), jednakże na ogół jest to najbardziej efektowna i rzucają się w oczy część kwiatu.
 |
| Źródło: Wikipedia. Schemat budowy okwiatu storczykowatych. Literą L oznaczono warżkę. |
 |
| Źródło: Wikipedia. Silnie wydęta warżka Paphiopedilum armeniacum. |
 |
| Źródło: Wikipedia. Warżka z ostrogą u Bulbophyllum echinolabium. |
 |
| Źródło: Wikipedia. Wyrostki na powierzchni trąbkowato zwiniętej warżki u Coelogyne tomentosa. |
 |
| Źródło: Wikipedia. Wypukły osadnik w kwiatostanie stokrotki pospolitej. |
11) Plewinka (łac. bracteola) - jest to łuseczka wyrastająca u podstawy kwiatów na dnie niektórych koszyczków u roślin z rodziny astrowatych. Jej morfologia ma duże znaczenie przy oznaczaniu niektórych gatunków roślin.
 |
| Źródło: Wikipedia. Z dna koszyczka rumiana polnego wyrastają żółte kwiaty rurkowate i błoniaste, wąskolancetowate plewinki. |
12) Spatha - jest to wyróżniający się liść przykwiatowy okrywający kwiatostan (podsadka). Spatha wyróżnia się kształtem, często jest także zabarwiona w sposób, który odróżnia się od innych liści. Ten typ liścia charakterystyczny jest dla niektórych jednoliściennych. Spatha występuje u palm czy obrazkowatych, u których ma kształt pochwiasty. U niektórych przedstawicieli rodzaju obrazki spatha pełni funkcję pułapkową.
 |
| Źródło: Wikipedia. Biała wewnątrz spatha czermieni błotnej. |
TYPY KWIATÓW:
 |
| Źródło: Wikipedia. Dwuwargowy kwiat Salvia patens. |
(a) - Kwiat grzbiecisty (łac. zygomorphicus) - jest to kwiat posiadający taką budowę i układ poszczególnych części okwiatu, że ma tylko jedną płaszczyznę symetrii. Kwiaty grzbieciste są bardziej zaawansowane ewolucyjnie, niż kwiaty promieniste posiadające dwie lub więcej osi symetrii. Występują np. w rodzinie bobowatych, storczykowatych, jasnotowatych i w wielu innych. Wśród kwiatów grzbiecistych istnieje wiele różnych form budowy. W rodzinie bobowatych mają specyficzną budowę; poszczególne płatki korony mają różny kształt i tworzą tzw. żagielek, łódeczkę i skrzydełka. W rodzinie storczykowatych jeden z płatków różni sie znacznie od pozostałych tworząc tzw. warżkę. U jasnotowatych zrośnięte płatki tworzą dwuwargową koronę. W rodzinie astrowatych są to tzw. kwiaty języczkowe. U niektórych kwiatów grzbiecistych występuje rurkowaty i zamknięty na końcu wyrostek - ostroga.
 |
| Źródło: Wikipedia. Zewnętrzne niby płatki słonecznika to w istocie pojedyncze kwiaty języczkowe. |
(b) - Kwiat języczkowy (łac. ligulatus, ang. ligular, ligulate) - jest to kwiat grzbiecisty, którego płatki korony zrośnięte są w dolnej części w rurkę, a w górnej części wyciągnięte w spłaszczony języczek. Kwiaty języczkowe występują np. w rodzinie astrowatych, wchodząc w skład kwiatostanu zwanego koszyczkiem. Koszyczek ten może składać się z samych kwiatów języczkowych, jak np. u mniszka, lub z samych kwiatów rurkowych. Najczęściej jednak w środku koszyczka znajdują się kwiaty rurkowe, zaś kwiaty języczkowe znajdują się na brzegu koszyczka. Zwykle kwiaty te są większe od kwiatów rurkowych wewnątrz koszyczka i barwniejsze, pełniąc rolę powabni. Często też są płone.
 |
| Źródło: Wikipedia. Schemat przedstawia budowę kwiatu języczkowe w rodzinie astrowatych. A - zalążnia, B - puch kielichowy; C - pręciki zrosłe w rurkę; D - płatki korony przekształcone w języczek; E - słupek. |
(c) Kwiat motylkowy (łac. papilionaceua) - jest to rodzaj kwiatu o specyficznej budowie występujący u roślin w rodzinie bobowatych (dawniej zwanych roślinami motylkowymi). Jest to kwiat grzbiecisty, którego kielich składa się z 5 zrośniętych działek, zaś korona z 5 zachodzących na siebie płatków o zróżnicowanej budowie. U większości gatunków są to płatki wolne (niezrośnięte), u nielicznych (np. koniczyny) płatki są zrośnięte. 2 dolne płatki tworzą tzw. łódeczkę, 2 boczne skrzydełka, a piaty wzniesiony jest do góry tworząc żagielek. Wewnątrz kwiatu jest jeden słupek i zwykle 10 pręcików, z których najczęściej dziewięć zrośniętych jest nitkami tworząc rurkę lub rynienkę, jeden pręcik zaś jest wolny. Do rynienki utworzonej przez zrośnięte nitki pręcików ścieka nektar. Pojedynczy słupek wyrastający wewnątrz rurki lub rynienki utworzonej z nitek pręcików ma przeważnie wydłużoną zalążnię i długą szyjkę. Jest to zwykle słupek jednokrotny i jednokomorowy, często lekko zagięty. Powstaje z niego owoc zwany strąkiem. Kwiaty motylkowe są przeważnie owadopylne, należą do zaawansowanych ewolucyjnie,a ich specyficzna budowa jest przystosowaniem do zapylenia krzyżowego. Tak np. u wyki płotowej pod ciężarem owada siadającego na łódeczce wysuwa się słupek, który pod znamieniem ma gęste włoski z przyklejonym pyłkiem z pręcików. Roślina jest przedprątna, nie może więc w tym czasie zapylić się własnym pyłkiem. Pyłek ten przykleja się do brzusznej strony owada i może być przez niego przeniesiony na inny kwiat wyki, w którym słupek jest już dojrzały.
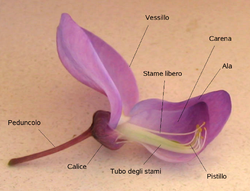
BUDOWA KWIATU MOTYLKOWEGO (WISTERII):
pedunculo - szypułka
vessillo - żagielek
calice - kielich
stame libero - słupek
pistillo - pręciki

(d) Kwiat paściowy - jest to kwiat na którym znajdują się urządzenia służące do przytrzymywania owadów zapylających tak, aby były zmuszone do zabrania pyłku. W przeciwieństwie do kwiatu pułapkowego owad znajduje się na zewnątrz kwiatu, przytrzymywany za odnóża lub aparat gębowy. W polskiej florze kwiaty paściowe wykształca np. ciemiężyk białokwiatowy. Ziarna pyłku każdego pylnika są ze sobą sklejone, tworząc pyłkowiny. Pomiędzy pylnikami są specjalne uczepki, które unieruchamiają owada na zasadzie zatrzasku. Owad, próbując się uwolnić, wyrywa pyłkowiny i przenosi na inny kwiat.
Po lewej stronie przedstawione zostały kwiaty paściowe ciemiężyka białokwiatowego (źródło: Wikipedia).
 |
| Źródło: Wikipedia. Ziarnopłon wiosenny. |
(e) Kwiat pełny - jest to określenie kwiatu ze zwielokrotnionymi listkami okwiatu. Kwiaty takie powstają w wyniku mutacji polegającej na modyfikacji części lub wszystkich pręcików oraz słupków i prowadzącej do rozwinięcia się ich w postaci listków okwiatu. Mutacja ta jest przykładem homeozji. W publikacjach specjalistycznych rośliny z kwiatami pełnymi oznaczane są skrótem fl. pl., którego rozwinięcie to łacińskie określenie flore pleno oznaczające "z pełnym kwiatem". Rośliny z kwiatami pełnymi często są cenionym walorem w przypadku roślin ozdobnych.
 |
| Źródło: Wikipedia. Kwiatostany męskie (kotki) u wierzby iwy. |
(f) - Kwiat pręcikowy, kwiat męski - jest to kwiat zawierający tylko pręciki, nieposiadający natomiast słupków. Oddzielne kwiaty męskie i kwiaty żeńskie występują u niektórych gatunków roślin nasiennych. Kwiaty pręcikowe mogą występować na jednej roślinie wspólnie z kwiatami słupkowymi, wówczas mówimy o jednopienności, a rośliny takie nazywamy roślinami jednopiennymi. Jeżeli w obrębie jednego gatunku na jednym osobniku występują wyłącznie kwiaty słupkowe, a na innym kwiaty pręcikowe, wówczas mówimy o dwupienności, a rośliny takie nazywamy roślinami dwupiennymi. Często kwiaty męskie tworzą kwiatostany. Kwiaty męskie wytwarzają pyłek. Aby mógł on dokonać zapylenia kwiatów żeńskich, musi zostać na nie przeniesiony, np. za pomocą wiatru (anemogamia), zwierząt (zoogamia) lub wody (hydrogamia). Po przekwitnięciu kwiaty pręcikowe obumierają. Występowanie oddzielnie kwiatów żeńskich i męskich (a zwłaszcza dwupienność), może być jednym z mechanizmów obronnych przed niekorzystnym dla większości roślin samozapyleniem.
 |
| Źródło: Wikipedia. Promienisty kwiat powojnika. |
(g) - Kwiat promienisty (łac. radiatus, actinomorphus) - jest to kwiat posiadający dwie lub więcej płaszczyzn symetrii. Jest to najstarszy ewolucyjnie typ budowy kwiatów. W rodzinie astrowatych jego odmianą są tzw. kwiaty rurkowe. Botanicy dla opisania budowy kwiatów promienistych używają narysu kwiatowego lub wzoru kwiatowego.
 |
| Źródło: Wikipedia. Promienisty kwiat Nymphea tetragona (Grzybienia karłowatego) |
- Kwiat rurkowy - jest to kwiat promienisty, mający dwie lub więcej płaszczyzn symetrii, o okwiecie rurkowatym lub lejkowatym. Ten typ kwiatów występuje np. w rodzinie astrowatych, wchodząc w skład kwiatostanu zwanego koszyczkiem. Koszyczek ten może składać się z samych kwiatów języczkowych, lub z samych kwiatów rurkowych, jak np. u ostów. Najczęściej jednak w środku koszyczka znajdują się kwiaty rurkowe, zaś kwiaty języczkowe znajdują się na brzegu koszyczka. Korona w kwiatach rurkowych jest zrosła, również zrośnięte pręciki tworzą rurkę wokół szyjki słupka. Po lewej stronie zostały przedstawione rurkowate kwiaty ostu zebrane w kwiatostan - koszyczek (źródło: Wikipedia).

SCHEMAT BUDOWY KWIATU RURKOWATEGO W RODZINIE ASTROWATYCH:
A - zalążnia, B - korna kwiatu, C - pręciki zrosłe w rurkę wokół słupka, D - szyjka i znamię słupka.

 |
| Źródło: Kwiaty pułapkowe kokornaku. |
Źródło: Wikipedia.
(h) Kwiat pułapkowy - jest to kwiat przystosowany do chwytania owadów i zatrzymywania ich w swoim wnętrzu tak długo, by dokonały zapylenia przyniesionym pyłkiem i zabrały dalej zebrany pyłek. Rośliny mające kwiaty pułapkowe to np. kokornak i obuwik.
(i) Kwiat słupkowy (kwiat żeński) - jest to kwiat zawierający tylko jeden lub więcej słupków, nieposiadający natomiast pręcików. Oddzielne kwiaty żeńskie i męskie występują u niektórych gatunków roślin nasiennych. Czasami kwiaty żeńskie tworzą kwiatostany. Kwiaty żeńskie mogą występować na jednej roślinie wspólnie z kwiatami męskimi, wówczas mówimy o jednopienności, a rośliny takie nazywamy roślinami jednopiennymi. Jeżeli w obrębie jednego gatunku na jednym osobniku występują wyłącznie kwiaty słupkowe, a na innym kwiaty pręcikowe, wówczas mówimy o dwupienności, a rośliny takie nazywamy roślinami dwupiennymi. Z kwiatów żeńskich powstaje po zapyleniu ich pyłkiem wytwarzanym przez kwiaty męskie nasiona i owoce. Pyłek może zostać przeniesiony przez wiatr (anemogamia), zwierzęta (zoogamia), wodę (hydrogamia0. Występowanie oddzielnie kwiatów żeńskich i męskich (a zwłaszcza dwupienność), może być jednym z mechanizmów obronnych przed niekorzystnym dla większości roślin samozapyleniem.
*WZÓR KWIATOWY* - wzór kwiatowy to zapis służący do przedstawiania budowy morfologicznej kwiatu za pomocą symboli, liter oraz liczb. Informuje on o liczbie elementów wposzczególnych okółkach kwiatu i o ich zrastaniu się, a także o położeniu dna kwiatowego względem zalążni. Stosowane są następujące oznaczenia:
P - KWIAT POJEDYNCZY, TJ. NIEZRÓŻNICOWANY NA KIELICH ORAZ KORONĘ (perigonium, perianthium) E - KIELISZEK (epicalyx)
K - KIELICH (calyx) C - KORONA (corolla) A - PRĘCIKOWIE (androeceum)
G - SŁUPKOWIE (gynaeceum)
Liczby umieszczane w indeksie dolnym po poszczególnych literach oznaczają liczbę elementów w okółku, np. C5 oznacza koronę o 5 płatkach, K3 oznacza kielich o 3 działkach. Jeżeli elementów danego typu jest więcej niż 12, to zamiast podawania ich liczby (która jest najczęściej wówczas zmienna nawet w różnych kwiatach tej samej rośliny), pisze się znak nieskończoności (), który czytamy wiele albo liczne. Obecnie panuje tendencja do zastępowania tego znaku literą "n".
*WZÓR KWIATOWY* - wzór kwiatowy to zapis służący do przedstawiania budowy morfologicznej kwiatu za pomocą symboli, liter oraz liczb. Informuje on o liczbie elementów wposzczególnych okółkach kwiatu i o ich zrastaniu się, a także o położeniu dna kwiatowego względem zalążni. Stosowane są następujące oznaczenia:
P - KWIAT POJEDYNCZY, TJ. NIEZRÓŻNICOWANY NA KIELICH ORAZ KORONĘ (perigonium, perianthium) E - KIELISZEK (epicalyx)
K - KIELICH (calyx) C - KORONA (corolla) A - PRĘCIKOWIE (androeceum)
G - SŁUPKOWIE (gynaeceum)
Liczby umieszczane w indeksie dolnym po poszczególnych literach oznaczają liczbę elementów w okółku, np. C5 oznacza koronę o 5 płatkach, K3 oznacza kielich o 3 działkach. Jeżeli elementów danego typu jest więcej niż 12, to zamiast podawania ich liczby (która jest najczęściej wówczas zmienna nawet w różnych kwiatach tej samej rośliny), pisze się znak nieskończoności (), który czytamy wiele albo liczne. Obecnie panuje tendencja do zastępowania tego znaku literą "n".

SYMBOLE:
+ - wskazuje, że elementy danego typu (płatki, pręciki) występują w dwóch okółkach, np. A5+5 oznacza pręciki w dwóch okółkach, w każdym po 5 pręcików.
( ) - oznacza zrastanie się (częściowe lub całkowite) elementów należących do tego samego okółka, np. C(5) wskazuje, że koronę tworzy 5 zrośniętych ze sobą płatków; A(9)+1 oznacza, że z 10 pręcików 9 zrosło się w rurkę (swymi nitkami), a dziesiąty jest wolny (=niezrośnięty).
[ ] - stosuje się, aby wskazać przyrastanie nitek pręcików do korony, w zapisie typu: [C(5)A5].
- - kreska nad liczbą słupków (lub owocolistków w przypadku słupkowia zrosłego) oznacza, że słupek jest dolny (a tym samym kwiat jest górny), pod tą liczbą - oznacza że słupek jest górny (a kwiat dolny). Kreska ta symbolizuje więc jak gdyby "poziom", na którym znajduje się dno kwiatowe. Np. oznacza słupek dolny (=zalążnię dolną) zbudowaną z 5 zrośniętych owocolistków.

Symbole symetrii kwiatu (umieszczane na początku wzoru kwiatowego):
- * – symetria promienista kwiatu
- ↓ – symetria grzbiecista (rzadko spotyka się znak ↑).
Wzory kwiatowe można zapisać dla każdego gatunku rośliny kwiatowej, w większości przypadków są jednak stałe dla całej rodziny lub podrodziny, a więc dla wielu blisko spokrewnionych rodzajów roślin.
KWIATOSTANY:
Kwiatostan (ang. inflorescence, łac. inflorescentium) - to wyodrębniające się od pozostałej części rośliny skupienie rozgałęzień pędów zakończonych kwiatami. W obrębie kwiatostanu znajdować się mogą poza nimi także liście przykwiatowe (podkwiatek i przysadka), a sam kwiatostan bywa wsparty liściem zwanym podsadką. Chociaż w typowej formie kwiatostan wykształcił się u roślin okrytozalążkowych, w języku polskim mianem kwiatostanów określa isę także szyszki żeńskie (makrostrobile) jak i szyszki męskie (mikrostrobile) roślin nagozalążkowych. U roślin okrytozalążkowych wyróżnia się dwa podstawowe typy kwiatostanów: groniaste i wierzchotkowate, różniące się sposobem rozgałęzienia.
KWIATOSTANY GRONIASTE: Posiadają one wyraźną oś główną rozgałęziającą się jednoosiowo (monopodialnie), rosnącą i wydająćą kolejne odgałęzienia boczne. W kwiatostanach groniastych kwiaty rozwijają się od nasady w kierunku wierzchołka, czyli od zewnętrznych części kwiatostanu ku środkowi. Najstarsze kwiaty znajdują się w efekcie u podstawy albo w zewnętrznych częsciach kwiatostanu, zaś namłodsze na szczycie lub w środku rozgałęzień. We wszystkich kwiatostanach groniastych oś główna przewyższa osie boczne. W kwiatostanach groniastych wyróżnia się:
PROSTE:
 |
| Źródło: Wikipedia. Kwiaty konwalii zebrane w grono. |
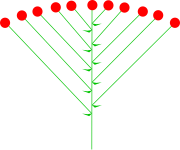
1) GRONO - ten typ kwiatostanu składa się z osi głównej, od której odchodzą osie boczne, które tworzą kwiaty szypułkowe. Przykładem grona jest kwiatostan konwalii. Występujące u niektórych gatunków listki pod każdym kwiatkiem to przysadki. Kwiaty w gronie mogą wyrastać na wszystkie strny, może być grono dwustronne, lub jednostronne (np. u mieczyka dachówkowatego). Grono może być też częścią kwiatostanu złożonego, gdy zamiast pojedynczych kwiatów na szypułkach z osi głównej wyrastają kwiatostany, np. koszyczki (jak u lepiężnika, czy kłoski). Potocznie mianem grona nazywa się także kiść winogron.
 |
| Źródło: Wikipedia. Baldachogrono rutewki orkolistnej. |
 |
| Źródło: Wikipedia. |
 3) KŁOS (łac. spica) - jest to rodzaj kwiatostanu groniastego, w którym na osadce siedzą kwiaty bezszypułkowe. Odmianą kłosa jest kwiatostan zwany kotką (np. u wierzb) i kolba (np. kwiatostany żeńskie kukurydzy).
3) KŁOS (łac. spica) - jest to rodzaj kwiatostanu groniastego, w którym na osadce siedzą kwiaty bezszypułkowe. Odmianą kłosa jest kwiatostan zwany kotką (np. u wierzb) i kolba (np. kwiatostany żeńskie kukurydzy). |
| Źródło: tinypic.com - Typy zbóż. |
 |
| Źródło: Wikipedia. Dwa rodzaje kotek: groniasta oraz kłosowa. |
- Kotka, bazia (łac. amentum) - odmiana kłosa lub grona, typ kwiatostanu, w któ®ym pojedyncze kwiaty osadzone są na osi pędu bez szypułek lub z krótkimi szypułkami. Od kłosa i grona różni się tym, że po spełnieniu swojej roli odpada w całości. Kotki mają przeważnie wiotką, zwieszającą się oś główną. Kotka należy do grupy kwiatostanó∑ otwartych, bez pączka szczytowego, czyli teoretycznie o nieograniczonym wzroście. Kwiaty zaczynają się rozwijać od nasady głównej osi, stopniowo w kierunku wierzchołka kotki. Najstarsze kwiaty znajdują się więc u nasady kotki. Należy także do grupy kwiatostanów groniastych, czyli kwiatostanów posiadających jedną główną oś. Kotki występują np. u wierzby, leszczyny. Występujące u podstawy każdego kwiatka w kotce listki to przysadki, a u podstawy całej kotki - podsadki. Ich występowanie, kształt, kolor i morfologia ma znaczenie przy oznaczaniu np. licznych gatunków wierzb, podobnie, jak wielkość i morfologia samej kotki.
 |
| Źródło: Wikipedia. Kotki męskie leszczyny pospolitej. |
 |
| Źródło: Wikipedia. Kolba u pałki szerokolistnej. |
- Kolba (łac. spadix) - jest to rodzaj kwiatostanu roślin zwany z rosyjskiego kaczanem. W botanice określeniem tym nazywa się kłos o silnie zgrubiałej osi. Zwykle w kolbie kwiaty ułożone są bardzo gęsto, ściśle obok siebie i są siedzące. Kolba występuje m.in. u niektórych roślin z rodziny arekowatych, obrazkowatych, wiechlinowatych (np. u kukurydzy zwyczajnej). Nierzadko w górnej części kolby kwiatów płodnych brak, znajdują się w niej natomiast komórki wydzielające substancje aromatyczne wabiące zapylaczy.

 |
| Źródło: Wikipedia. Baldach czosnku południowego. |
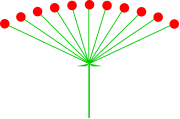 4) BALDACH (łac. umbella) - jest to u roślin rodzaj kwiatostanu, w którym pojedyncze kwiaty wyrastają na mniej więcej jednakowej długości szypułkach z jednego miejsca na szczycie pędu. Najwcześniej rozkwitają kwiaty zewnętrzne, czym bliżej środka tym później. Baldach, zwany też baldachem prostym należy do grupy kwiatostanów groniastych o silnie skróconej osi głównej. Ten typ kwiatostanu, występuje np. u czosnku, pierwiosnka. Odmianą baldachu prostego jest podbaldach, u którego wszystkie szypułki dorastają do tego samego poziomu. Czasami baldach prosty wchodzi w skład kwiatostanów złożonych, np. baldachu złożonego. Wyrastające u niektórych roślin listki pod baldachem noszą nazwę pokrywek.
4) BALDACH (łac. umbella) - jest to u roślin rodzaj kwiatostanu, w którym pojedyncze kwiaty wyrastają na mniej więcej jednakowej długości szypułkach z jednego miejsca na szczycie pędu. Najwcześniej rozkwitają kwiaty zewnętrzne, czym bliżej środka tym później. Baldach, zwany też baldachem prostym należy do grupy kwiatostanów groniastych o silnie skróconej osi głównej. Ten typ kwiatostanu, występuje np. u czosnku, pierwiosnka. Odmianą baldachu prostego jest podbaldach, u którego wszystkie szypułki dorastają do tego samego poziomu. Czasami baldach prosty wchodzi w skład kwiatostanów złożonych, np. baldachu złożonego. Wyrastające u niektórych roślin listki pod baldachem noszą nazwę pokrywek.- BALDASZEK (łac. umbellula) - jest to mały baldach, najczęściej będący częścią składową kwiatostanu zwanego baldachem złożonym. W baldaszku pojedyncze kwiaty wyrastają na mniej więcej jednakowej długości szypułkach z jednego miejsca na osi pędu. Najwcześniej rozkwitają kwiaty zewnętrzne, czym bliżej środka tym później. Baldaszki są charakterystyczną częścią składową kwiatostanów roślin z rodziny selerowatych (dawniej zwanych baldaszkowymi). Czasami występują też jako części składowe innych kwiatostanów, np. u konwalijki dwulistnej kilkukwiatowe baldaszki wchodzą w skłąd grona. Listki występujące czasami u nasady baldaszka noszą nazwę pokrywek.
 |
| Źródło: Wikipedia. Baldaszki u kopru ogrodowego. |
 |
| Źródło: Wikipedia. Duże pokrywki u przewiercienia okrągłolistnego. |
 |
| Źródło: Wikipedia. Drobne pokrywki u Heracleum lanatum. |
 |
| Źródło: Wikipedia. Główka. |
 |
| Źródło: Wikipedia. Główka koniczyny. |
 6) KOSZYCZEK (łac. antbodium, calathidium) - jest to typ kwiatostanu groniastego. Na szerokim dnie kwiatowym gęsto osadzone są bezszypułkowe, zwykle drobne kwiaty. Mogą to być same rurkowate kwiaty (jak np. u ostów) lub wyłącznie kwiaty języczkowe (jak np. u mleczka i mniszka). Najczęściej jednak na zewnątrz, jako kwiaty brzeżne umieszczone są kwiaty języczkowe, a wewnątrz kwiaty rurkowate (jak np. u słonecznika). Zewnętrzne kwiaty brzeżne pełnią wówczas zwykle rolę powabni, mającej za zadania zwabianie owadów, są duże i barwne, często też bezpłodne. Kwiaty w koszyczku z zewnątrz otoczone sa łuskami okrywy. Często kielich kwiatów przekształcony jest w puch kielichowy. U podstawy poszczególnych kwiatów w koszyczku często znajduje się łuseczka zwana plewinką, będąca przekształconą przysadką. Koszyczek występuje u roślin z rodziny astrowatych (dawniej nazywanych złożonymi). Należy do grupy kwiatostanów otwartych, w których ciągle rozwijają się nowe kwiaty. Najstarsze kwiaty znajdują się na zewnętrznych częściach koszyczka, czym bliżej jego środka, tym młodsze kwiaty. Jest zaawansowanym ewolucyjnie typem kwiatostanu. Często, gdy roślina wytwarza wiele koszyczków, tworzą one kwiatostan zbiorowy, mogą być zebrane, podobnie, jak pojedyncze kwiaty np. w grono, wiechę, itd.
6) KOSZYCZEK (łac. antbodium, calathidium) - jest to typ kwiatostanu groniastego. Na szerokim dnie kwiatowym gęsto osadzone są bezszypułkowe, zwykle drobne kwiaty. Mogą to być same rurkowate kwiaty (jak np. u ostów) lub wyłącznie kwiaty języczkowe (jak np. u mleczka i mniszka). Najczęściej jednak na zewnątrz, jako kwiaty brzeżne umieszczone są kwiaty języczkowe, a wewnątrz kwiaty rurkowate (jak np. u słonecznika). Zewnętrzne kwiaty brzeżne pełnią wówczas zwykle rolę powabni, mającej za zadania zwabianie owadów, są duże i barwne, często też bezpłodne. Kwiaty w koszyczku z zewnątrz otoczone sa łuskami okrywy. Często kielich kwiatów przekształcony jest w puch kielichowy. U podstawy poszczególnych kwiatów w koszyczku często znajduje się łuseczka zwana plewinką, będąca przekształconą przysadką. Koszyczek występuje u roślin z rodziny astrowatych (dawniej nazywanych złożonymi). Należy do grupy kwiatostanów otwartych, w których ciągle rozwijają się nowe kwiaty. Najstarsze kwiaty znajdują się na zewnętrznych częściach koszyczka, czym bliżej jego środka, tym młodsze kwiaty. Jest zaawansowanym ewolucyjnie typem kwiatostanu. Często, gdy roślina wytwarza wiele koszyczków, tworzą one kwiatostan zbiorowy, mogą być zebrane, podobnie, jak pojedyncze kwiaty np. w grono, wiechę, itd.  |
| Źródło: Wikipedia. Koszyczek słonecznika - na zewnątrz duże kwiaty języczkowe wyglądające jak płatki pojedynczego kwiatu, wewnątrz zaś drobne kwiaty rurkowate. |
 |
| Źródło: Wikipedia. kłosy pszenicy formy ościstej. |
- 1) KŁOS ZŁOŻONY (łac. spica composita) - rodzaj kwiatostanu groniastego, w którym na osadce siedzą kwiaty bezszypułkowe. Odmianą kłosa jest kwiatostan zwany kotką (np. u wierzb) i kolba (np. kwiatostany żeńskie kukurydzy). Czasami zamiast pojedynczych kwiatów na osi głównej występują kłoski, taki kwiatostan nazywa się kłosem złożonym (łac. spica composita). Występuje on u wielu gatunków roślin z rodziny wiechlinowatych, np. jęczmienia, pszenicy, żyta, bliźniczki psiej trawki. Kłos może być też częścią składową innych kwiatostanów, np. kukurydzy kwiaty męskie tworzące kłoski zebrane są w wiechę złożoną.
 |
| Źródło: Wikipedia. Budowa kłoska: 1 - plewa; 2- plewka dolna; 3 - ość plewki; 4 - plewka górna; 6 - pręciki. |
- KŁOSEK (łac. spicula) - jest to kwiatostan charakterystyczny dla traw, okryty od zewnątrz dwiema plewami [plewa, z łac. glumae - to u roślin, głównie u gatunków z rodziny wiechlinowatych, niewielki, łódeczkowatego kształtu, zielony lub bezzieleniowy liść (podsadka) wyrastający u podstawy kłosków. Przeważnie występują dwie plewy: plewa dolna (łac. gluma interior) i plewa górna (gluma superior), rzadziej jedna (np. u życicy) lub więcej niż dwie (np. 4 u mozgi). U niektórych gatunków plewy są większe od plewek i obejmują kłosek (np. owies), u innych są mniejsze od plewek i tylko podpierają kłoski (np. żyto)], obejmujący jeden lub więcej (najczęściej 2-6) kwiatów, z których każdy znajduje się w okrywie łuskowatych plewek. Kłoski nie występują pojedynczo, lecz wchodzą w skład innych kwiatostanów złożonych - kłosów, gron i wiech. Kłoski mogą być drobne (np. u mietlic) lub osiągać do kilku cm długości (np. u owsa). Widoczne są jako zbite, kulistawe lub częściej w różnym stopniu wydłużone części składowe kwiatostanów złożonych.
 |
| Źródło: Wikipedia. Schemat kłoska dwukwiatowego: 1 - plewa, 2 - plewa dolna, 3 - ość plewki, 4 - plewka górna, 5 - łuszczka, 6 - pręciki, 7 - słupek. |
*Łuszczka (łac. lodiculae) - to u gatunków z rodziny wiechlinowatych drobny delikatny listek wewnętrznego okółka okwiatu. W okresie kwitnienia łuszczki pęcznieją rozchylając plewki i ułatwiając wydostanie się na zewnątrz pręcików. Większość gatunków posiada dwie łuszczki w kwiecie, wyjątkowo występuje jedna (mietlica) lub trzy (ostnica, bambus, ryż). Łuszczek nie mają gatunki, których plewki podczas kwitnienia nie rozchylają się (np. wyczyniec, tomka). Według najpopularniejszej teorii łuszczki są przekształconymi dwoma listkami wewnętrznego okółka listków okwiatu w typowym kwiecie jednoliściennych. trzeci, tylny listek uległ redukcji. Inna teoria wyprowadza obecny kwiat traw od kwiatu bezokwiatowego. Według tej teorii dwie łuszczki to podzielony liść przykwiatowy, ewentualna trzecia łuszczka to kolejny liść przykwiatowy.
 |
| Źródło: Wikipedia. Łuszczki (g) na schemacie budowy kłoska traw: a1, a2 - plewy; b - plewka dolna; c - plewka górna; d - pręciki; e - znamię słupka; f - zalążnia słupka; g - łuszczki. |
*Plewka - jest to niewielki łuseczkowaty listek występujący głównie w kwiatach roślin z rodziny wiechlinowatych. U niektórych gatunków plewki zakończone są długą ością. Najczęściej występują dwie plewki:
- PLEWKA DOLNA (łac. lemma) - wyrasta ona u nasady kwiatu i jest odpowiednikiem przysadki (a więc nie jest elementem kwiatu traw). Może być wypukła - z widoczną linią grzbietową lub zaokrąglona - albo prawie płaska. Posiada na szczycie ość lub jest bezostna, może być owłiosiona lub naga, tępa lub zaostrzona, z kilka (do 10) nerwami.
- PLEWKA GÓRNA (łac. palea) - zwykle mniejsza od plewki dolnej, wyrasta nieco wyżej po przeciwległej stronie osi kwiatu. Powstaje ona z zewnętrznego okółka okwiatu, zredukowanego do dwóch listków (stąd zwykle jest dwunerwowa). Plewki górnej nie posiadają tylko nieliczne gatunki, np. wyczyniec. Po krzekwitnięciu kwiatu plewki zwykle twardnieją. U niektórych gatunków, np. u owsa plewki podczas dorjzewania zrastają się z ziarniakiem tworząc jego dodatkową okrywę (powstaje tzw. ziarniak oplewiony), u innych, np. u pszenicy czy żyta są związane z ziarniakiem bardzo luźno i łatwo odpadają podczas omłotu zboża lub samorzutnie. Wymłocone plewki i plewy stanowią paszę objętościową suchą.
 |
| Źródło: Wikipedia. Wiecha owsa. |
 2) WIECHA - jest to typ kwiatostanu groniastego złożonego, w którym na głównej osi pędu wykształcają się boczne odgałęzienia drugiego i trzeciego rzędu, a na nich wyrastają kwiaty. Istnieje jeszcze tzw. wiecha złożona, w której rozgałęzienia nie są zakończone pojedynczymi kwiatami, lecz kwiatostanami - np. kłoskami (m.in. u licznych gatunków z rodziny wiechlinowatych, np. owies, męskie kwiatostany kukurydzy) lub koszyczkami (u niektórych przedstawicieli astrowatych, np. lepiężnik, nawłoć).
2) WIECHA - jest to typ kwiatostanu groniastego złożonego, w którym na głównej osi pędu wykształcają się boczne odgałęzienia drugiego i trzeciego rzędu, a na nich wyrastają kwiaty. Istnieje jeszcze tzw. wiecha złożona, w której rozgałęzienia nie są zakończone pojedynczymi kwiatami, lecz kwiatostanami - np. kłoskami (m.in. u licznych gatunków z rodziny wiechlinowatych, np. owies, męskie kwiatostany kukurydzy) lub koszyczkami (u niektórych przedstawicieli astrowatych, np. lepiężnik, nawłoć).  |
| Źródło: Wikipedia. Baldachy złożone u barszczu Mantegazziego. |
3) BALDACH ZŁOŻONY (łac. umbella composita) - jest to rodzaj kwiatostanu, w którym na szypułkach baldachów I rzędu wyrastają baldachy II rzędu (zwane baldaszkami). Czasami spotyka się baldachy podwójnie złożone, w których występują baldaszki drugiego rzędu (np. u barszczu Mantegazziego). Kwiatostan jest wówczas trzypiętrowy. W poszczególnych baldachach najwcześniej rozkwitają kwiaty zewnątrzne, czym bliżej środka, tym później. Baldach złożony należy do grupy kwiatostanów groniastych o silnie skróconej osi głównej. Ten typ kwiatostanu jest bardzo charakterystyczny zwłaszcza dla roślin z rodziny selerowatych (dawniej zwanych baldaszkowatymi), np. kopru, marchwi. Wyrastające u niektórych roślin listki u nasady baldacha złożonego to pokrywy.
*Pokrywy (łac. bracteae involucri) - to rodzaj podsadek (podsadka to w botanice liść, z kąta którego wyrasta kwiatostan lub część kwiatostanu). Pokrywy wyrastają u nasady baldacha złożonego w rodzinie selerowatych (baldaszkowatych). Występowanie pokryw, ich morfologia i wielkość ma znaczenie przy oznaczaniu niektórych gatunków tych roślin.
 |
| Źródło: Wikipedia. Podzielone pokrywy u marchwi zwyczajnej. |
KWIATOSTANY WIERZCHOTKOWE: posiadają one kwiat na szczycie osi, który kończy jej wzrost na długość (zamknięte), rozgałęziają się sympodialnie. Gdy oś główna zakończy wzrost - rozwija się jedno lub więcej odgałęzień bocznych, każde z kwiatem szczytowym kończącym jego wzrost (rośliny, które wytwarzają tylko pojedynczy kwiat na szczycie pędu kończący jego wzrost nazywane są haplokaulicznymi). Kwiaty kwitną kolejno od środka, tj. od szczytu osi głównej, do coraz dalszych od niego odgałęzień bocznych. Kwiatostany wierzchotkowe dzieli się na:
- wierzchotkę jednoramienną (monochazjum) - z pojedynczymi rozgałęzieniami osi na kolejnych rzędach:
- 1) SIERPIK (łac. drepanium) - rodzaj kwiatostanu z grupy wierzchotek jednoramiennych. Wszystkie kwiaty wyrastają na szypułkach po jednej tylko stronie głównego pędu. Gdy patrzymy na kwiatostan z góry widać, że ułożone one są w jednej linii. W sierpiku kwiat wyrasta na szczycie pędu kończąc jego wzrost. Gdy główny pęd zakończy wzrost, zaczynają się rozwijać pędy boczne, każdy z kwiatem na szczycie. Kwiaty w sierpiku kwitną poczynając od środka, na zewnątrz. Ten typ kwiatostanu występuje np. u żywokostu, niezapominajki. Bardzo podobny do sierpika jest skrętek, który różni się tylko tym, że widziany z góry ma kwiaty uożone nie w linii prostej, lecz zygzakowato.
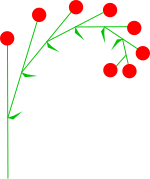
 |
| Źródło: Sierpik żywokostu lekarskiego. |
 |
| Źródło: Wikipedia. Skrętek. |
- 3) Dwurzędka - jest to rodzaj kwiatostanu z grupy wierzchotek jednoramiennych. Ma kwiat na szczycie pędu. Gdy kwiat ten zaczyna się rozwijać, wyrasta (zazwyczaj z kąta liścia) odgałęzienie drugiego rzędu, które znowu kończy się kwiatem. Kolejne odgałęzienia wyrastają naprzemiennie po obu stronach. Jest to dość rzadko spotykane typ kwiatostanu, występuje np. u niektórych gatunków z rodziny kosaćcowatych.
- Dwurzędka jest bardzo podobna do wachlarzyka, różni się od niego jednak tym, że szypułki poszczególnych kwiatów mają mniej więcej tę samą długość i wskutek tego kwiaty w dwurzędce nie znajdują się na jednym poziomie. Kwiaty w dwurzędce zaczynają kwitnąć od kwiatu znajdującego się w początkowej części kwiatostanu, stopniowo w kierunku pojawiających się osi drugiego, trzeciego i następnych rzędów. Jeżeli kwiatostan jest wzniesiony, kwiaty kwitną od dołu w górę. Listki, z kątów których wyrastają te osie noszą nazwę podsadek.
- 4) Wachlarzyk - jest to rodzaj kwiatostanu z grupy wierzchotek jednoramiennych. Ma kwiat na szczycie pędu. Od osi tego pędu wyrasta (zazwyczaj z kąt aliścia) odgałęzienie drugiego rzędu, które znowu kończy się kwiatem. Kolejne odgałęzienia wyrastają naprzemiennie po obu stronach. Wachlarzyk jest bardzo podobny do dwurzędki - różni się od niej tym, że wszystkie kwiaty dorastają mniej więcej do tego samego poziomu. Najdłuższe szypułki mają kwiaty dolne, czym wyżej w górę, tym krótsze. Kwiaty w wachlarzyku zaczynają kwitnąć od kwiatu znajdującego się w początkowej części kwiatostanu, stopniowo w kierunku pojawiąjących się osi drugiego, trzeciego i następnych rzędów, czyli od zewnątrz do wewnątrz kwiatostanu. Listki, z kątów których wyrastają te osie noszą nazwę podsadek.
 |
| Źródło: Wikipedia. Dwurzędka. |
 |
| Źródło: Wikipedia. Wachlarzyk. |

- 6) Wierzchotka wieloramienna, wielopromienista (pleiochazjum) - wytwarza w podobny sposób jak wierzchotka dwuramienna nie dwa, lecz kilka kwiatów (wilczomlecz).

- 7) Wiecha z wykształconym kwiatem szczytowym, który kończy wzrost (np. winorośl, lilak):
 |
| Źródło: Wikipedia. Podbaldach z owocami u kaliny koralowej. |
- - Podbaldach (łac. corymbothyrsus) - rodzaj kwiatostanu, będącego odmianą wiechy, w której kwiaty dorastają mniej więcej do tej samej wysokości. Swoim wyglądem natomiast przypomina baldachogrono lub baldach złożony. Podbaldach należy do grupy kwiatostanów wierzchotkowych. Na końcu osi pędu głównego znajduje się kwiat, który kończy wzrost pędu. Gdy pęd główny przestaje rosnąć, zaczynają się rozwijać pędy boczne. Kwiatostany takie nazywa się zamkniętymi. Podbaldach występuje np. u kaliny, centurii pospolitej. U niektórych gatunków w rodzinie astrowatych na szczycie osi zamiast pojedynczych kwiatów występują kwiatostany - koszyczki. Powstaje więc kwiatostan złożony - podbaldach z koszyczkami, jak np. u krwawnika.
 |
| Źródło: Wikipedia. Podbaldach u Viburnum globosum. |
- 8) Kłębik - gęsty i drobny kwiatostan wierzchotkowy o skróconej osi i krótkoszypułkowych lub siedzących kwiatach typowy dla komosowatych. Skupione kłębiki tworzą wiechowate i kłosokształtne kwiatostany złożone.
 |
| Źródło: Wikipedia. Kłębik. |
 |
| Źródło: Wikipedia. Skupione kłębiki tworzące kwiatostan złożony u komosy ryżowej. |
9) Pęczek - jest to kwiatostan wierzchotkowy charakteryzujący się bardzo skróconą osią (podobnie jak przy kłębiku) oraz kwiatami osadzonymi na nieco dłuższych szypułkach. Taki rodzaj kwiatostanu występuje między innymi u wiązów (Ulmus L) czy u niektórych gatunków z rodziny rdestowatych (Polygonaceae Juss.).
 |
| Źródło: Wikipedia. Pęczek - kwiatostan wiązu górskiego (Ulmus glabra). |
SPECYFICZNE MODYFIKACJE: Kwiatostany w różnych grupach systematycznych roślin ulegają daleko idącej modyfikacji trudnej do ujęcia w klasycznej typologii morfologicznej. Wyróżnia się m.in. kwiatostany złożone zawierające na szczycie osi głównej kwiatostan główny (pierwotny), a na bocznych - kwiatostany boczne (określane jako pędy powtarzające - parakladia). U neiktórych roślin pnących (np. u Ophiocaulon cissampeloides i Bowiea volubilis) kwiatostany ulegają modyfikacjom zmieniającym je w organy przypominające wąsy czepne - kwiaty ulegają redukcji, szypuły sztywnieją i stają się hakowate. W rodzinie obrazkowatych u wielu przedstawicieli występują kwiatostany pułapkowe. U wielu roślin poza kwiatami w obrębie kwiatostanów powstają bulwki służące do rozmnażania wegetatywnego. Specyficzne kwiatostany powstają w niektórych rodzinach, np. u wilczomleczowatych kwiaty zebrane są w cyjacjum. U figowców osadnik tworzy dzbanuszkowatą otoczkę wokół kwiatów (ang. hepanthodium). U wielu roślin kwiatostan wykształca się w sposób przypominający pojedynczy kwiat i określany jest wówczas pseudancjum.
1) Cyjacjum - struktura zawierająca organy generatywne występująca u roślin wilczomleczowatych (Euphorbieae), szczególny rodzaj kwiatostanu. Taki kwiatostan funkcjonuje jak jeden obupłciowy kwiat, czyli jest rodzajem pseudanthium. Rozbieżności w interpretacji, czy struktura jest kwiatem, czy kwiatostanem sięgają XVIII wieku. Jako kwiat traktował strukture Linneusz w opisie z 1753 rku, jako kwiatostan opisał Lamarck w roku 1786. Chociaż w kolejnych latach opis jako kwiatostan pojawiał się częściej, problem interpretacji pozostaje otwarty. Specyfika struktury jest na tyle duża, że ani określenie kwiatostan ani kwiat nie w pełni zadowala botaników. Cyjacjum ma kształt miseczki z wypustkami. Kwiatostan zawiera jeden, centralnie umieszczony słupek, stanowiący kwiat żeński, otoczony przez cztery lub pięć grup pręcików, stanowiących kwiaty męskie. Struktura zawiera często miodniki oraz efektowne podsadki. Cyanthia zwykle zebrane są w kwiatostany wyższych rzędów, wierzchotki.
 |
| Źródło: Wikipedia. Cyjacjum Euphorbia tridentata.
2) Pseudancjum - jest to kwiatostan podobny z wyglądu do pojedynczego kwiatu. Termin ten dotyczy kwiatostanów w przypadku których poszczególne kwiaty je tworzące są drobne i rosną skupione, np. w formie główki lub koszyczka. Dodatkowo listki okrywy lub brzeżne kwiaty języczkowate otaczają cały kwiatostan tworząc odpowiednik korony. Funkcjonalnie kwiatostany takie funkcjonują jak pojedynczy kwiat. Do przykładów pseudancjów należą koszyczki astrowatych, kolby obrazkowatych i cyjacja wilczomleczowatych. W budowie kwiatostanów występują także postaci podobne do pseudanjów, w których skupione kwiatostany otoczone są powabnią powstającą z liści przykwiatowych, jak u jarzmianki Astrantia czy dereni Cornus. W morfologicznej klasyfikacji typów kwiatów pseudancja traktowane są jak pojedyncze kwiaty i tak kolbowate kwiatostany obrazkowatych łączone są z kwiatami pułapkowymi i paściowymi, a cyjacja wilczomleczów zaliczane są do "kwiatów talerzykowatych". Nazwa "pseudancjum" stosowana jest także w odniesieniu do teorii, według której obupłciowy kwiat typowy dla okrytozalążkowych miał wyewoluować z kwiatostanu złożonego z kwiatów rozdzielnopłciowych.
|
Brak komentarzy:
Prześlij komentarz